


Beoordeling van cellulaire opname-efficiëntie volgens meerdere remmers van Fe3O4-Au Core-Shell-nanodeeltjes:mogelijkheid om specifieke endocytose in colorectale kankercellen te beheersen
Abstract
Magnetiet (Fe3 O4 )-goud (Au) kern-schil nanodeeltjes (NP's) hebben unieke magnetische en optische eigenschappen. In combinatie met biologische groepen kunnen deze NP's nieuwe strategieën bieden voor biomedische toepassingen, zoals medicijnafgifte en kankertargeting. Hier presenteren we een effectieve methode voor de controleerbare cellulaire opname van magnetische core-shell NP-systemen in combinatie met biologische groepen. Van Vimentine, het structurele eiwit, is biochemisch bevestigd dat het fagocytose krachtig beïnvloedt. Bovendien beïnvloedt vimentine de internalisatie van exogene materialen in cellen, hoewel onder meerdere remmingen van biologische delen. In deze studie demonstreren we de cellulaire internalisatieprestaties van Fe3 O4 -Au core-shell NP's met oppervlaktemodificatie met behulp van een combinatie van biologische groepen. De fotofluorescentie van met vimentine gemerkte NP's bleef onaangetast onder meerdere remmingstests, wat aangeeft dat de NP's minimaal werden beïnvloed door nystatine, dynasore, cytochalasine D en zelfs het Muc1-antilichaam (Ab). Bijgevolg geeft dit resultaat aan dat de Mucl Ab zich kan richten op specifieke moleculen en specifieke endocytose kan beheersen. Bovendien tonen we de mogelijkheid aan om specifieke endocytose in colorectale kankercellen te beheersen.
Inleiding
Nanomaterialen hebben nieuwe wegen geopend voor klinische diagnostiek en therapieën. Vooral nanodeeltjes (NP's) zijn een van de belangrijkste hulpmiddelen en zijn gebruikt in toepassingen zoals biosensoren [1, 2], diagnostiek [3, 4] en gerichte medicijnafgiftesystemen [5, 6]. Voor biomedische toepassingen bestaan NP's over het algemeen uit organische materialen die het oppervlak op kernmaterialen omringen [7,8,9]. De kernmaterialen, die bestaan uit magnetische materialen, halfgeleidermaterialen of andere soorten materialen, hebben nuttige fysisch-chemische eigenschappen en het buitenste organische oppervlak biedt chemische stabiliteit en functionaliteit aan de NP's. Voor toepassingen in biologische targetingsystemen zijn niet alleen de fysisch-chemische eigenschappen, maar ook het buitenste organische oppervlak, die biogefunctionaliseerd zijn voor targeting, kritische parameters. Voorbeelden van targeting-eenheden voor functionalisering zijn antilichamen of liganden die specifiek zijn voor een target. Afhankelijk van de biogefunctionaliseerde materialen op het buitenoppervlak, worden de endocytosemechanismen van NP's bepaald. Het mechanisme waarmee NP's cellen kunnen binnendringen, is het onderwerp geweest van veel recent onderzoek vanwege hun belang in nanogeneeskundetoepassingen [10,11,12,13,14,15].
In het bijzonder zijn magnetische NP's op grote schaal gebruikt in veel specifieke toepassingen voor locatietargeting, waaronder celsortering [16, 17], MRI [18], DNA-isolatie [19], medicijnafgifte [20], hyperthermiebehandeling [21] en kanker richten [22]. Van de verschillende magnetische NP's zijn magnetiet-nanokristallen het meest gebruikt in biomedische toepassingen vanwege hun biocompatibiliteit en chemische stabiliteit. Hoewel er veel inspanningen zijn geleverd voor biomedische toepassingen met behulp van magnetische NP's, zijn er nog steeds enkele kritieke problemen, zoals een goede dispergeerbaarheid in waterige oplossing, functionaliteit en biocompatibiliteit. Om deze problemen te overwinnen, hebben veel onderzoeken zich gericht op de oppervlaktemodificatie van NP's met behulp van een verscheidenheid aan functionele groepen (bijv. Carboxyl- en aminegroepen) [23]. De hechting van de functionele groepen aan het oppervlak van magnetiet NP's is echter een tijdrovend en arbeidsintensief proces. Gezien dit feit zijn Au-gecoate magnetische NP's van het kern-shell-type aantrekkelijk omdat het Au-oppervlak gemakkelijk kan worden gekoppeld aan biomoleculen en organische materialen.
Vooral de magnetische eigenschappen van de magnetische kern-Au shell NP's maken de magnetische scheiding mogelijk, verhogen de resolutie in MRI-beeldvorming en kunnen worden toegepast op hyperthermietherapie. Bovendien zijn de superieure chemische bindingseigenschappen van goud voordelig voor het bouwen van receptor-gemedieerde leveringssystemen voor specifieke kankertargeting [24,25,26].
In de afgelopen decennia hebben veel onderzoekers receptor-gemedieerde toedieningssystemen voor kankertargeting gerapporteerd [27,28,29].
Receptor-gemedieerde targeting van kankercellen is een vorm van actieve targeting. De keuze van het doelwit is de sleutel voor effectieve actieve targeting en de doelwitten moeten tot overexpressie worden gebracht op het extracellulaire membraan. De meeste onderzoekers hebben monoklonale antilichamen gebruikt voor de behandeling van kanker, en het therapeutische effect zou sterk kunnen worden verhoogd wanneer monoklonale antilichaamtherapieën worden gecombineerd met conventionele chemotherapie [30]. Ondanks het succes van monoklonale antilichaamtherapie, vertonen monoklonale antilichamen verschillende beperkingen bij het richten op kanker. Hun grote omvang (ongeveer 150 kDa) is een belangrijk obstakel voor tumorpenetratie [31, 32], en hun lage stabiliteit en lage oplosbaarheid belemmeren hun wijdverbreide gebruik [33]. De inhomogene richting van hun hechting op de targeting carrier wordt ook beschouwd als een obstakel voor niet-specifieke binding. Om antilichamen met verbeterde tumorpenetratie te produceren, is een breed scala aan antilichaamformaten ontwikkeld en getest [34]. Naast klassieke antilichamen is er een uniek antilichaamformaat aanwezig in soorten uit de familie Camelidae. De zogenaamde zware keten antilichamen (HCAbs) komen van nature voor in het perifere bloed en de melk van deze soorten. De antigeenbindende fragmenten van dergelijke HCAb's zijn samengesteld uit één enkel domein, het variabele domein van de zware keten (VH) van het kameelachtige HCAb (VHH). Het VHH, recombinant verkregen na klonering en expressie in bacteriën of schimmels, wordt een nanobody genoemd. Het heeft een molecuulgewicht van 11-15 kDa en is het kleinste antilichaam van alle mAbs [35,36,37]. Niet alleen hun kleine formaat maakt ze potentieel geschikt als targeting-probes tegen antigenen op geïsoleerde locaties, maar ook hun gemakkelijk aanpasbare terminale uiteinde is aantrekkelijk voor toepassing bij targeting op kanker.
De efficiënte levering van NP's met geschikte targeting en internalisatie van cellen zijn ook belangrijke factoren in het leveringssysteem. Er is gemeld dat vimentine een belangrijke rol speelt als onderdeel van de aanhechting van pathogenen en intracellulaire toegangswegen. Het stilleggen van de genexpressie van vimentine remt fagocytose [38], terwijl gesplitst vimentine een signaal is dat fagocytose significant verhoogt [39]. Daarom is het neutraliseren van de resistentie tegen celfagocytose veroorzaakt door vimentine op het celoppervlak belangrijk voor een efficiënte afgifte van nanodeeltjes.
In deze studie onderzoeken we de endocytoseroutes van nanobody-gelabeld Fe3 O4 -Au core-shell NP's gemodificeerd met PEG (polyethyleenglycol) spacers met verschillende lengtes. Vimentine, waarvan bekend is dat het een sterk effect heeft op fagocytose door biochemische experimenten [39], werd vergeleken als een controle en er werd bevestigd dat het effectief inwerkt op celinternalisatie van NP's. Bovendien wordt Muc1, dat een glycoproteïne op het celoppervlak is en tot overexpressie wordt gebracht in verschillende kankers, zoals pancreas-, borst-, long- en maagkanker, gebruikt als een kankergerichte biomarker. We hebben de efficiënte internalisatie van Fe3 . bevestigd O4 Au core-shell NP's en de methoden voor controleerbare targeting op kankercellen via de Muc1-receptor-gemedieerde endocytose-route in coloncellen.
Materialen en methoden
Materialen
Goud(III)acetaat (Au(OOCCH3)3, 99,9%) werd verkregen van Alfa Aesar. Andere chemicaliën, waaronder ijzer (III) acetylacetonaat (Fe(acac)3 , 99,9%), 1,2-hexadecaandiol (C14H29CH(OH)CH2(OH), 90%), poly(ethyleenglycol)-blok-poly(propyleenglycol)-blok-poly(ethyleenglycol) (PEG-PPG- PEG) en octylether (C8H17OC8H17, 99%) werden gekocht bij Sigma-Aldrich en gebruikt zoals ontvangen. Alfa-pyridyl-2-disulfid-omega-carboxysuccinimidylester poly(ethyleenglycol) (OPSS-PEG-NHS) (2K, 5K en 10K) werd gekocht bij Nanocs. Natriumbicarbonaat, WST-1, chloorpromazine, nystatine, cytochalasine D, dynasore, brefeldine A (BFA), monensine en trypanblauw werden gekocht bij Sigma-Aldrich. Cy3 en Cy7.5 werden gekocht bij Lumiprobe. Anti-Muc1 Ab werd gekocht bij Abcam Inc. (Cambridge, MA). Fosfaatgebufferde zoutoplossing (PBS), Dulbecco's gemodificeerde Eagle's medium en foetaal runderserum werden gekocht bij Invitrogen Corp.
Synthese van Fe3 O4 -Au Core-Shell NP's
De Fe3 O4 -Au core-shell NP's werden gesynthetiseerd via een nano-emulsiemethode. Het syntheseproces voor kern-schil NP's bestaat uit twee stappen:(1) vorming van de Fe3 O4 kern-NP's en (2) coating van de Au-schaal op de magnetische NP's. In de eerste stap, de Fe3 O4 NP's werden bereid uit een gemengde oplossing van Fe(acac)3 (0,1766 g of 0,5 mmol), 1,2-hexadecandiol (0,6468 g of 2,5 mmol) en blokcopolymeer (poly(ethyleenoxide)-poly(propyleenoxide)-poly(ethyleenoxide); PEO-PPO-PEO) ( 0,4 ~ 1,2 g) in octylether. De gemengde oplossing werd verwarmd tot 300°C om de Fe-precursor te verminderen. De vorming van Fe3 O4 kern NP's werd voltooid door de verwarmde oplossing af te koelen. Het tweede proces werd continu uitgevoerd zonder enig zuiveringsproces na de vorming van de magnetische kern. Au-precursoren (0,2338 g of 0,62 mmol) en 1,2-hexadecandiol (0,88 g, 3,4 mmol) werden toegevoegd aan de emulsie bestaande uit Fe3 O4 NP's, en vervolgens werd de gemengde oplossing verwarmd tot 230°C. Na afkoeling tot kamertemperatuur werd de emulsie geprecipiteerd door centrifugatie en werden de kern-schil NP's gescheiden.
Constructie van recombinante anti-Muc1-VHH 5-24 K10-expressievector
Polymerasekettingreactie (PCR) werd uitgevoerd met behulp van de voorwaartse primer 5'-CCGAATTCGCCGATGTGCAGCTGACCGAG-3' en de omgekeerde primer 5'-CGG CTCGAGCTTCTTCTTCTTCTTCTTCTTCTTCTTCTTGCCTGAGGAGACGGTGACCTG-3'. Het PCR-product werd verteerd met EcoRI en XhoI en met gel gezuiverd met behulp van de QIA quick Gel Extraction Kit (QIAGEN, Valencia, CA, VS). Het gezuiverde PCR-product werd gekloneerd in met EcoRI/XhoI gedigereerd pET-23a (Novagen, Darmstadt, Duitsland). Escherichia coli (E. coli ) DH5a (RBC Bioscience, Xindian, Taiwan) werd getransformeerd met het resulterende construct door hitteschok en geselecteerd op LB-agarplaten die 100 g/mL ampicilline bevatten (Duchefa Biochemie, Haarlem, Nederland).
Expressie en zuivering van recombinant eiwit
Om recombinant anti-Muc1-VHH 5-24 K10-eiwit tot expressie te brengen en te zuiveren, heeft E. coli BL21-stammen (RBC Bioscience, Xindian, Taiwan) werden getransformeerd met pET-23a-anti-Muc1-VHH 5-24 K10. Bacteriën werden vervolgens gekweekt in LB-bouillon met ampicilline (100 g / ml). Eiwitexpressie werd geïnduceerd door isopropyl β-d-thiogalactoside (IPTG) (Duchefa Biochemie, Haarlem, Nederland) bij een eindconcentratie van 0,4 mM gedurende 5 h bij 37 °C. Bacteriële pellets werden opnieuw gesuspendeerd in lysisbuffer (50 mM NaH2 PO4 , pH 8,0; 300 mM NaCl) gevolgd door sonicatie op ijs gedurende 10 min. Sonicated lysaten werden gecentrifugeerd bij 20.000 × g gedurende 20 min bij 4 °C en onderworpen aan Ni-NTA His·Bind Resin (Peptron, Daejeon, Korea). His-gelabelde eiwitten die aan de hars waren gebonden, werden geëlueerd met elutiebuffer (50 mM NaH2 PO4 , pH 8,0; 300 uM NaCl; 150 mM imidazool). Gezuiverd eiwit werd gescheiden op 15% SDS-PAGE-gel.
Wijziging van Core-Shell Fe3 O4 -Au NP's
OPSS-PEG-NHS op verschillende lengtes (2, 5 en 10 K) werd opgelost in 0,1 M natriumbicarbonaat voor activering van de thiolgroepen. Geactiveerde OPSS-PEG-NHS werd toegevoegd aan de oplossing van gesynthetiseerde core-shell Fe3 O4 -Au NP's en gedurende 12 uur geroerd bij 4 °C. De thiolgroepen van geactiveerd OPSS-PEG-NHS waren covalent gekoppeld aan het Au-oppervlak van de core-shell NP's. Vervolgens werd een nanobody-oplossing (0,25 mg/mL) toegevoegd aan het gePEGyleerde Fe3 O4 -Au core-shell NP's gedurende 12 h bij 4 °C. De aminegroepen van de tien lysine (K)-staarten aan het uiteinde waren covalent gekoppeld aan de NHS-groepen van OPSS-PEG-NHS bij pH 8.3. Cy3 en Cy7.5 werden gelabeld aan de resterende aminegroepen van het nanobody.
Internalisatiecurve
CT26-cellen werden gezaaid bij 5 × 10 3 cellen per putje in een plaat met 96 putjes met heldere bodem en gedurende 24 u bij 37 °C in 250 L kweekmedium geïncubeerd in 5% CO2 in het donker. Het medium werd verwijderd en 250 l vers kweekmedium met 50 μg/ml Cy3-gelabeld Fe3 O4 -Au NP's en PEG-Cy3 of PEG-nanobody-Cy3-gelabelde NP's werden aan elk putje toegevoegd. De cellen werden verder geïncubeerd gedurende verschillende perioden (0, 10, 20, 30, 60, 120 en 360 min). De cellen werden vervolgens driemaal gewassen met PBS om vrije NP's te verwijderen, en de fluorescentie van elk putje werd gemeten met trypanblauw als een membraan-ondoordringbare fluorescentiedoder door SpectraMAX GEMINI (Molecular Devices, CA, VS). Elk experiment werd uitgevoerd met gelijke hoeveelheden NP's (50 g/ml) en drie keer herhaald [40].
Remmingstest
CT26-cellen werden gezaaid bij 5 × 10 3 cellen per putje in een plaat met 96 putjes met heldere bodem en gedurende 24 u bij 37 °C in 250 L medium geïncubeerd in 5% CO2 in het donker. Het medium werd verwijderd en 250 l vers kweekmedium met 20 g/ml chloorpromazine (CPZ), 50 g/ml nystatine, 20 g/ml cytochalasine D, 25 g/ml dynasore, 20 g/ml BFA, 140 μg/ ml monensin of 5 M anti-Muc1 Ab werd toegevoegd en de cellen werden gedurende 1 uur geïncubeerd.
Het medium werd opnieuw verwijderd en 250 L kweekmedium met 50 μg/ml Cy3-gelabeld Fe3 O4 -Au NP's, PEG-Cy3-gelabelde NP's, PEG-nanobody-Cy3-gelabelde NP's of vimentin-Cy3-gelabelde NP's zijn toegevoegd.
Na 1 uur bij 37 °C en 5% CO2 , de cellen werden drie keer gewassen met PBS om vrije NP's te verwijderen, en de fluorescentie werd gemeten met trypanblauw als een membraan-ondoordringbare fluorescentiedover door SpectraMAX GEMINI (Molecular Devices, CA, VS). Elk experiment werd uitgevoerd met gelijke hoeveelheden NP's (50 g/ml) en vier keer herhaald.
Resultaten en discussie
De core-shell NP's werden gesynthetiseerd met een gepubliceerde methode [16, 17]. Transmissie-elektronenmicroscopie (TEM)-waarnemingen in Fig. 1a, b laten zien dat de Fe3 O4 -Au core-shell NP's waren bolvormig met een gemiddelde diameter van 13,5 nm en een smalle grootteverdeling.
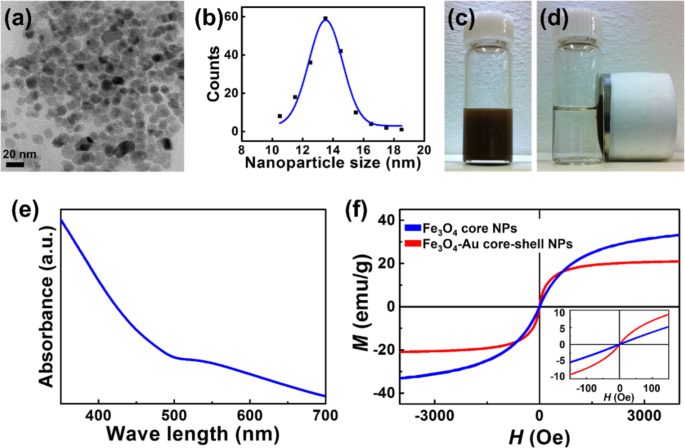
Karakterisering van het gesynthetiseerde Fe3 O4 -Au core-shell NP's. een , b TEM-waarnemingen van het gesynthetiseerde Fe3 O4 -Au core-shell NP's. c , d NP's in waterige oplossing, voor en na het aanleggen van een extern magnetisch veld. e UV-absorptiepiek van gesynthetiseerde core-shell NP's verschijnt bij ~ 530 nm. v Magnetische hysteresislussen van Fe3 O4 kern
De toename van de ~ 8.5 nm van de kern NP (Fe3 O4 ) komt voort uit de coating van ~ -2,5 nm dikke Au-schaal op het kernoppervlak, wat resulteert in een kern-schaal NP. Een TEM-beeld met hoge resolutie met de snelle Fourier-transformatie (FFT)-analyse van de Fe3 O4 -Au core-shell NP is opgenomen in aanvullende informatie Fig. S1.
Het product vervaardigd in organisch oplosmiddel werd gezuiverd met behulp van magnetische scheiding en overgebracht naar het water.
De kern-schil NP's waren goed gedispergeerd en stabiel in water zonder enige oppervlaktemodificatie, dankzij resterende blokcopolymeren die op de NP's aanwezig waren.
Figuur 1c en d tonen de NP's in waterige oplossing voor en na het aanleggen van een extern magnetisch veld. Onder een extern magnetisch veld veranderden de kern-schil NP's snel van een homogene dispersie (figuur 1c) in een heldere en transparante oplossing (figuur 1d).
De absorptieband van de kern-schil NP's werd onderzocht met behulp van UV-Vis-spectrometrie. Zoals getoond in Fig. 1e, verscheen een absorptiepiek bij ~ 530 nm, wat de aanwezigheid van Au op het oppervlak van de NP's aangeeft (aanvullende informatie Fig. S2 bevat het resultaat van EDX-gegevens voor Fe3 O4 -Au core-shell NP's). Omdat het monster was gezuiverd, toonden de optische resultaten de vorming van de kern-schaalstructuur aan.
Magnetische hysteresislussen werden verkregen uit vibrerende monstermetingen om de magnetische eigenschappen van de Fe3 te onderzoeken O4 core en de core-shell NP's. Beide NP's vertoonden superparamagnetisch gedrag met een coërciviteit van bijna 0 Oe bij kamertemperatuur (Fig. 1f).
Zoals gerapporteerd in eerdere werken, was de gevoeligheid van kern-schil NP's hoger dan die van de magnetiet NP's, wat deels te wijten zou kunnen zijn aan nabijheidseffecten en unieke ruimtelijke configuraties [41, 42]. Bovendien zijn de verzadigingsmagnetisaties van de kern-NP's en de kern-schil-NP's respectievelijk ~ 37 emu/g en ~ 21 emu/g bij 10 kOe. Het verschil in de Ms komt voort uit het bestaan van een niet-magnetische component (Au) in de core-shell NP's.
Het VHH 5-24 K10-gen werd in-frame gekloneerd om pET-23a-anti-Muc1-VHH 5-24 K10 te produceren na PCR-amplificatie (Fig. 2a). Het recombinante eiwit werd tot expressie gebracht in E. coli BL21 dat werd getransformeerd met pET-23a-anti-Muc1-VHH 5-24 K10 na inductie met IPTG en gezuiverd door Ni-NTA His-Bind-hars. Recombinant anti-Muc1-VHH 5-24 K10 werd gemakkelijk tot expressie gebracht in E. coli als een oplosbaar 18-kDa-eiwit. Uit een kweek van 1 L verkregen we 1 ± 0,5 mg gezuiverd recombinant anti-Muc1-VHH 5-24 K10.
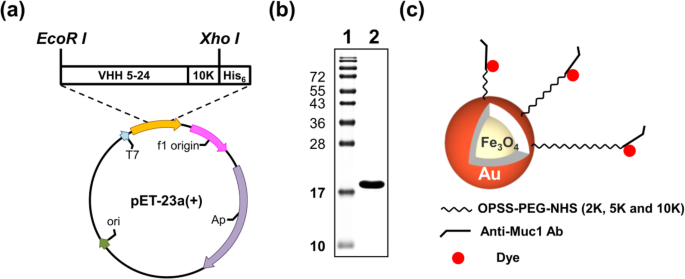
Expressie en zuivering van anti-Muc1-VHH 5-24 K10-fusie-eiwit. Het VHH 5-24 K10-gen werd in-frame gekloond om a . te produceren pET-23a-anti-Muc1-VHH 5-24 K10 na b zuivering van Mucl-VHH 5-24 K10-fusie-eiwit. Gezuiverd eiwit werd gescheiden op 15% SDS-PAGE. Baan 1 eiwitladder. Baan 2 gezuiverd eiwit. c Schematische illustratie van PEG-nanobody-dye-gelabelde NP's die in dit onderzoek zijn gebruikt
Het gezuiverde eiwit werd geverifieerd met 15% SDS-PAGE-gel. Coomassie-blauwkleuring van het gezuiverde eiwit onthulde dat het> 95% zuiver was (Fig. 2b). De gesynthetiseerde Fe3 O4 -Au NP's werden in drie stappen gemodificeerd, namelijk PEGylatie, antilichaam-tagging en kleurstoflabeling (Fig. 2c). Na elke wijzigingsstap werd de zeta-potentiaal gemeten om de succesvolle wijziging te bevestigen. Tabel 1 toont het effect van modificatie op de overeenkomstige zetapotentialen. Het zeta-potentieel van kale kern-schil NP's was − 19,8 ± 6,68 mV. PEGylering van NP's werd uitgevoerd met behulp van OPSS-PEG-NHS. Om een reeks nanocomplexen met verschillende groottes te produceren, werd OPSS-PEG-NHS met verschillende lengtes (2 K, 5 K en 10 K) gebruikt.
Na PEGylatie waren de zeta-potentialen verminderd (− 44,9 ± 8,19 mV, − 40,7 ± 7,88 mV en − 39,6 ± 8,74 mV voor respectievelijk 2 K, 5 K en 10 K).
Interessant is dat na nanobody-tagging de zeta-potentialen duidelijk toenam (− 38,5 ± 5,61 mV, − 23,3 ± 8,61 mV en − 31,8 ± 7,37 mV voor respectievelijk 2 K, 5 K en 10 K).
Na kleurstofmarkering namen de zeta-potentialen ook toe (− 12,5 ± 7,25 mV, − 17,7 ± 3,94 mV en − 10,6 ± 4,72 mV voor respectievelijk 2, 5 en 10 K).
Het zeta-potentieel van kale Fe3 O4 -Au core-shell NP's was − 19,8 ± 6,68 mV. Na PEGylatie werd de zeta-potentiaal verlaagd tot bijna − 40 mV. Deze resultaten geven aan dat de PEG-moleculen goed covalent gebonden waren aan de Au-schaal van de kern-schil-NP's omdat PEG-moleculen negatief geladen N-hydroxysuccinimide-functionele groepen hebben. Ondertussen nam het zeta-potentieel toe na kleurstofmarkering op het nanobody (− 38,5 ± 5,61 mV voor NP-PEG2 K-nanobody en − 12,5 ± 7,25 mV voor NP-PEG2 K-nanobody-kleurstof). Deze uitkomst is redelijk omdat het recombinante nanobody tien lysine-staarten heeft aan het uiteinde. Elk type nanobody werd gecategoriseerd door zeta-potentiaalmeting, en om de antilichaambinding op het nanobody te bepalen, hebben we de fluorescentie voor elk nanobody-type gemeten.
Zoals getoond in Fig. 3, hebben we bevestigd dat alle soorten nanodeeltjes en nanobodies een goede cellulaire opname en internalisatie hebben in afwezigheid van remmerbeperkingen. Een cellulaire internalisatiecurve werd verkregen uit cellen die waren geïncubeerd in aanwezigheid van 50 g/ml Cy3-gelabeld Fe3 O4 -Au NP's, PEG-Cy3-gelabelde NP's en PEG-nanobody-Cy3-gelabelde NP's voor verschillende perioden (tussen 0 en 360 min) na het verwijderen van media, het uitwassen van vrije NP's en tenslotte het meten van de totale fluorescentie van de cellen met trypan blauw (Fig. 3).
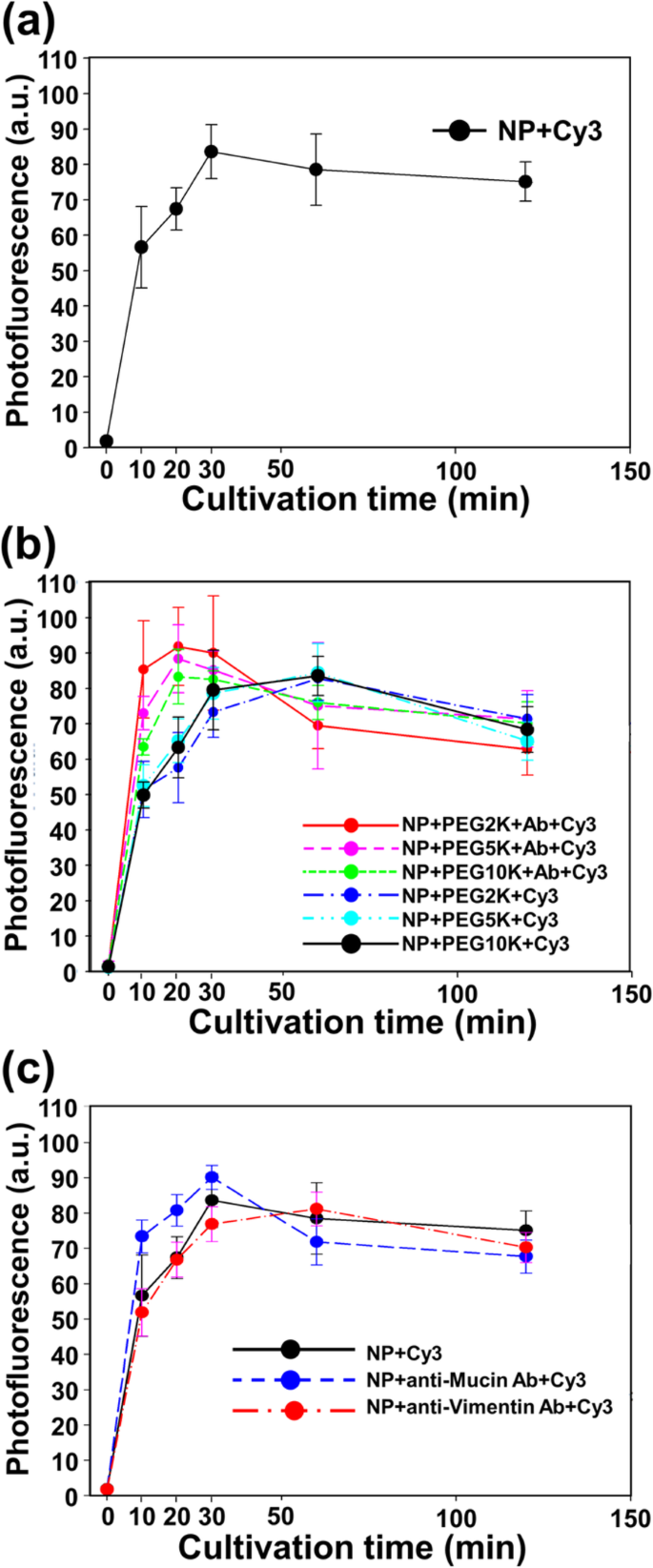
Genormaliseerde fotofluorescentie van CT26-mucinecellen na incubatie met 50 μg/mL a Fe3 O4 -Au NP's, b PEGylated NP's en PEG-nanobody-tagged NP's, en c Fe3 O4 -Au NP's, nanobody-gelabelde NP's en vimentine-gelabelde NP's bij 37 ° C in 5% CO2 voor verschillende perioden (10, 20, 30, 60, 120 en 360 min)
Volgens het resultaat van de meting van de fluorescentie-intensiteit, kunnen we bepalen dat de NP's binnen 1 uur in de cellen werden geïnternaliseerd (figuur 3a). De fluorescentie-intensiteit van NP bereikte een maximum binnen 1 h, en de fluorescentie-intensiteit nam geleidelijk af na het bereiken van een stabiele toestand. Hoewel er een klein tijdsverschil is, afhankelijk van de aanwezigheid van Ab, was de fluorescentie-intensiteit per kweektijd niet significant verschillend van het resultaat van blote NP's (figuur 3b). Zoals getoond in Fig. 3c, werd bevestigd dat het effect van Ab, door gebruik te maken van heterogeen Ab van Mucl en vimentine, verwaarloosbaar was bij de cellulaire opname en internalisatie van NP.
Met WST-1-assay is de levensvatbaarheid (%) van CT26-mucinecellen afhankelijk van de concentratie en oppervlaktemodificatie van Fe3 O4 -Au core-shell NP's werden geschat na verschillende blootstellingstijden (Fig. 4a, b). De levensvatbaarheid van CT26-cellen vertoonde geen significante verschillen na 24 uur en 48 uur blootstelling, noch met variërende doses, noch na oppervlaktemodificatie van de NP's. De levensvatbaarheid van de cellen was groter dan 90% op zowel de kale Fe3 O4 -Au NP's (Fig. 4a) en de oppervlakte-gemodificeerde NP's (Fig. 4b).
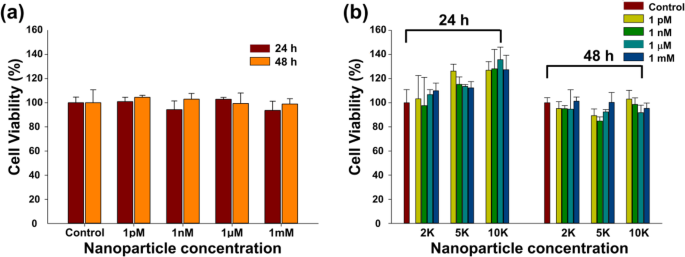
Levensvatbaarheid van CT26-mucinecellen behandeld met kale Fe3 O4 -Au core-shell NP's en oppervlakte-gemodificeerde Fe3 O4 -Au NP's in verschillende concentraties. een Fe3 O4 -Au NP's en b PEG-nanobody-Cy3-gelabelde NP's. Elke experimentele grafiek vertegenwoordigt het gemiddelde van een reeks van vier verschillende experimenten
Figuur 5 geeft aan dat NP's de CT26-mucinecellen binnengingen via verschillende endocytose-routes (clathrine-gemedieerde, caveolae-gemedieerde en fagocytose/macropinocytose-routes). Interessant is dat Fig. 5b laat zien dat anti-Muc1 Ab ook voornamelijk de endocytose van PEG-nanobody-Cy3-gelabelde NP's beïnvloedde. Om de route van NP-internalisatie te begrijpen, hebben we geprobeerd de endocytoseroutes te remmen met specifieke chemische remmers (Fig. 5). Het was algemeen bekend dat de routes van endocytose in drie typen kunnen worden verdeeld:clathrine-gemedieerde, caveolae-gemedieerde en macropinocytose/fagocytose.
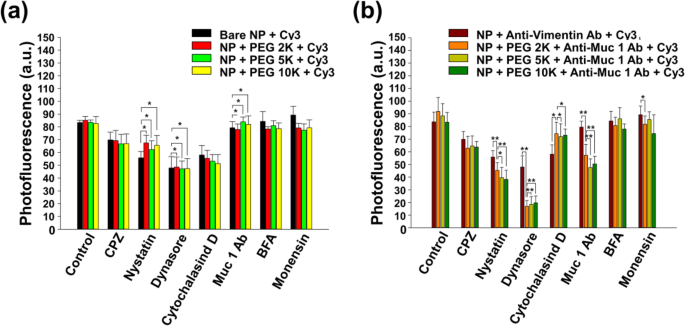
Genormaliseerde fotofluorescentie van CT26-mucinecellen behandeld met chemische endocytoseremmers gedurende 1 h en geïncubeerd met 50 μg/mL a kale core-shell NP's en b nanobody-gelabelde NP's bij 37 °C in 5% CO2 gedurende 30 min. Remmers met een statistisch effect op de internalisatie (Studenten t test, p (*) <0,05 en p (**) <0.01) zijn gemarkeerd met zwarte sterretjes
In deze studie werden remmers gebruikt als een eerste benadering om de internalisatie van nanobody-gelabelde NP's te onderzoeken. CPZ (clathrine-gemedieerde endocytoseremmer), nystatine (caveolae-gemedieerde endocytoseremmer), dynasore (dynamineremmer), cytochalasine D (fagocytose/macropinocytoseremmer), BFA (Golgi-apparaatvernietiger), monensin (lysosoomremmer) of anti-Muc1 Ab (receptor-/transporter-specifieke concurrent) werd gedurende 1 uur met cellen geïncubeerd. CPZ, nystatine, dynasore en cytochalasine D beïnvloedden de endocytose van NP's (Fig. 5a).
De targetinggroep is de sleutel tot het succes van kankertargeting, wat uitzonderlijk belangrijk is bij kankertherapie. Voor targeting is effectieve oppervlaktemodificatie erg belangrijk om de therapeutische efficiëntie te verhogen en bijwerkingen te beperken. Nanobody-gelabeld Fe3 O4 -Au core-shell NP's werden met succes gemaakt van de gesynthetiseerde NP's en recombinant nanobody. Tabel 1 laat duidelijk zien dat elke wijzigingsstap met succes werd uitgevoerd.
Levensvatbaarheid van de cellen is een van de essentiële elementen voor de biologische toepassing van nanomaterialen. De levensvatbaarheid van de cellen was groter dan 90% op de kale kern-schil NP's en gemodificeerde NP's (Fig. 4a, b). Deze resultaten impliceren dat kale Fe3 O4 -Au NP's en gemodificeerde NP's geen significante concentratie- en modificatie-afhankelijke cytotoxiciteit veroorzaakten en dat de gemodificeerde NP's geschikt waren voor biologische toepassing.
De onderzoeken naar internalisatie-efficiëntie en het remmende effect van NP's bieden belangrijke informatie om de mechanismen te begrijpen waarmee NP's cellen binnenkomen. GePEGyleerde NP's werden relatief langzaam geïnternaliseerd in cellen in vergelijking met kale NP's, maar nanobody-gelabelde NP's werden iets sneller in cellen geïnternaliseerd dan blote NP's (Fig. 3a, b). Omdat PEGylatie een bekende methode voor oppervlaktemodificatie is om de internalisatie van NP's te voorkomen, is de internalisatietendens van gePEGyleerde NP's gemakkelijk te verklaren. Bovendien induceerde het nanobody de endocytose van NP's. Om de specificiteit van het nanobody te controleren, hebben we de internalisatiesnelheid van vimentine Ab-gelabelde NP's bevestigd. Interessant is dat met vimentine Ab-gelabelde NP's de cellulaire internalisatie niet bevorderden (Fig. 3c).
Deze resultaten geven aan dat het nanobody effectief de internalisatie van nanomaterialen in CT26-mucinecellen kan induceren en impliceren dat de internalisatietendens kan worden gecontroleerd met specifieke modificatie van het buitenmembraan van NP's. Bovendien werden de mechanismen van de endocytose van nanobody-gelabelde NP's duidelijk aangetoond via remmingstesten en confocale microscopie-beeldvorming. De fotofluorescentie van zowel de nanobody-gelabelde NP's als de niet-gelabelde NP's vertoonden vergelijkbare afnemende waarden wanneer ze werden gekweekt met CPZ, nystatine of dynasore (Fig. 5a, b). CPZ, nystatine en dynasore spelen een rol bij het remmen van respectievelijk clathrine-gemedieerde endocytose, caveolae-gemedieerde endocytose en dynamin, een grote GTPase die betrokken is bij het ontluiken en splitsen van ontluikende blaasjes van oudermembranen. Daarbij namen de fotofluorescentiewaarden in beide gevallen snel af omdat dynamine nauw verwant is aan de productie van blaasjes voor clathrine-gemedieerde en caveolae-gemedieerde endocytose. Zoals getoond in Fig. 5a, vertoonden zowel niet-gelabelde (Fig. 5a) als nanobody-gelabelde NP's (Fig. 5b) snelle afnamen in fotofluorescentie.
In het bijzonder vertoonden de nanobody-gelabelde NP's significant lagere fotofluorescentiewaarden in CPZ, nystatine en dynasore. Bovendien bevestigden we dat de niet-gelabelde NP's sterker werden beïnvloed dan de nanobody-gelabelde NP's wanneer ze werden toegepast op cytochalasine D, een celpermeabel toxine dat de polymerisatie van actinefilamenten voor fagocytose blokkeert [43]. Deze resultaten impliceren dat niet-gelabelde NP's werden geïnternaliseerd via meerdere mechanismen, zoals clathrine-gemedieerde endocytose, caveolae-gemedieerde endocytose en fagocytose. Bijgevolg hangt de cellulaire internalisatie van nanobody-gelabelde NP's af van clathrine-gemedieerde en caveolae-gemedieerde endocytose. Bovendien was de hoeveelheid cellulaire opname van nanobody-gelabelde NP's aanzienlijk verminderd wanneer cellen werden gekweekt met het Muc1-antilichaam (figuur 5b). Dit resultaat geeft aan dat het vrije Muc1-antilichaam een rol speelt als concurrent van het nanobody op de gemodificeerde NP's bij het hechten aan het CT26-celmembraan en dat het Muc1-antilichaam een belangrijke rol speelt bij de celinternalisatie van de gemodificeerde NP's. Vreemd genoeg vertoonden vimentine Ab-gelabelde NP's duidelijke verschillen in vergelijking met nanobody-gelabelde NP's in termen van remmingsvermogen. De fotofluorescentie van vimentine-gelabelde NP's bleef onaangetast onder meerdere remmingstests, wat aangeeft dat de NP's minimaal werden beïnvloed door nystatine, dynasore, cytochalasine D en zelfs de Muc1 Ab. Dit fenomeen zou een bewijs kunnen zijn van de werkzaamheid van vimentine, waarvan biochemisch is bevestigd dat het fagocytose sterk beïnvloedt [39]. Bijgevolg geeft dit resultaat aan dat de Mucl Ab zich kan richten op specifieke moleculen en specifieke endocytose kan beheersen.
Zoals getoond in Fig. 6, werden analoge resultaten verkregen wanneer de cellen werden behandeld met Cy7.5-gelabelde kale kern-schil NP's en PEG-nanobody-gelabelde NP's, wat wijst op een vergelijkbare cellulaire opname in beide gevallen in afwezigheid van dynasore-remming. Dynasore-remming induceerde duidelijk lagere cellulaire internalisatie van de PEG-nanobody-gelabelde NP's in vergelijking met kale NP's (Fig. 6b, onderste rij). Deze resultaten impliceren dat er twee endocytosemechanismen zijn, namelijk niet-specifieke endocytose van kale NP's en beperkte endocytose van nanobody-gelabelde NP's via het dynaminmolecuul. Zodra het nanobody hecht aan het externe celmembraan, kunnen de nanobody-gelabelde NP's gemakkelijk door het celmembraan gaan vanwege de gelijktijdige activering van de clathrin- en caveolae-gemedieerde mechanismen. Bijgevolg zouden we kunnen veronderstellen dat de belangrijkste mechanismen zowel door clathrin als door caveolae gemedieerde endocytose zijn voor de internalisatie van nanobody-gelabelde NP's in CT26-mucinecellen.
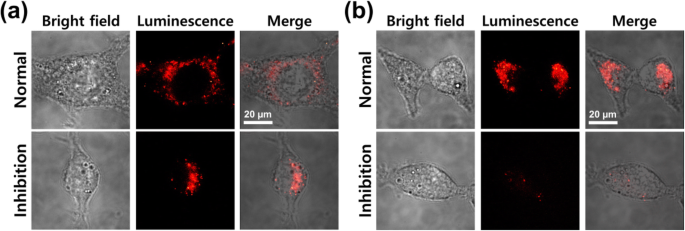
Confocale microscopische beeldvorming van CT26-mucinecellen geïncubeerd met a Fe3 O4 -Au NP's en b PEG-nanobody-gelabelde NP's gedurende 1 uur bij 37 °C in 5% CO2 incubator, voor en na dynasore-remming (1 ng/mL)
Conclusies
Nanomaterialen voor kankertargeting en controleerbare insertie van exogene materialen zoals medicijnen, genen en peptiden zijn cruciale vooruitgang in biomedische toepassingen. Deze bekende maar creatieve concepten kunnen strategieën bieden voor nieuwe therapeutische methoden. In dit artikel hebben we een verbeterde cellulaire opname van Fe3 . aangetoond O4 -Au core-shell NP's na PEGylatie met het Muc1-antilichaam. De belangrijkste endocytosemechanismen van nanobody-gelabelde NP's werden aangetoond, wat de mogelijkheid aantoont van controleerbare specifieke endocytose in colorectale kankercellen. Deze bevindingen geven inzicht in de targeting tussen nanobody-tagged NP's en colorectale kankercellen om het ontwerp van zeer efficiënte targeting-dragers te helpen.
Beschikbaarheid van gegevens en materialen
Alle gegevens die tijdens dit onderzoek zijn gegenereerd of geanalyseerd, zijn opgenomen in dit gepubliceerde artikel.
Afkortingen
- Fe3 O4 :
-
Magnetiet
- Au:
-
Goud
- NP's:
-
Nanodeeltjes
- Ab:
-
Antilichaam
- HCAbs:
-
Zware-keten antilichamen
- VHH:
-
VH van de kameelachtige HCab
- PEG:
-
Poly(ethyleenglycol)
- PPG:
-
Poly(propyleenglycol)
- PCR:
-
Polymerase kettingreactie
- TEM:
-
Transmissie-elektronenmicroscopie
- PEO-PPO-PEO:
-
Poly(ethyleenoxide)-poly(propyleenoxide)-poly(ethyleenoxide)
- OPSS-PEG-NHS:
-
Orthopyridyldisulfide PDP PEG succinimidylester
- BFA:
-
Brefeldin A
- CPZ:
-
Chloorpromazine
Nanomaterialen
- Hoogrendement grafeen zonnecellen
- Nanodeeltjes voor kankertherapie:huidige vooruitgang en uitdagingen
- Op cellen gebaseerde medicijnafgifte voor kankertoepassingen
- Nanotechnieken inactiveren kankerstamcellen
- Endotheelcellen targeten met multifunctionele GaN/Fe-nanodeeltjes
- Gemodificeerd hypervertakte polyglycerol als dispergeermiddel voor groottecontrole en stabilisatie van gouden nanodeeltjes in koolwaterstoffen
- Invloed van Ag-nanodeeltjes met verschillende afmetingen en concentraties ingebed in een TiO2-compacte laag op de conversie-efficiëntie van perovskiet-zonnecellen
- Antiproliferatief en apoptose-triggerend potentieel van op paclitaxel gebaseerde gerichte lipide nanodeeltjes met verbeterde cellulaire internalisatie door transferrinereceptoren - een onderzoek in le…
- Groeiende gouden nanostructuren voor vormselectieve cellulaire opname
- Het apoptose-effect op leverkankercellen van gouden nanodeeltjes gemodificeerd met lithocholzuur
- Door zirkoniumoxide veroorzaakte toxische effecten in osteoblastachtige 3T3-E1-cellen