


F-actine reguleert osteoblastische differentiatie van mesenchymale stamcellen op TiO2-nanobuisjes via MKL1 en YAP/TAZ
Abstract
Titanium en titaniumlegeringen worden veel gebruikt in orthopedische implantaten. Het wijzigen van de nanotopografie biedt een nieuwe strategie om de osseo-integratie van titaniumsubstraten te verbeteren. Filamenteuze actine (F-actine) polymerisatie, als een mechanische belastingsstructuur, wordt algemeen beschouwd als betrokken bij celmigratie, endocytose, celdeling en celvormbehoud. Of F-actine erbij betrokken is en hoe het functioneert bij door nanobuisjes geïnduceerde osteogene differentiatie van mesenchymale stamcellen (MSC's) moet nog worden opgehelderd. In deze studie hebben we TiO2 . gefabriceerd nanobuisjes op het oppervlak van een titaniumsubstraat door anodische oxidatie en hun kenmerken gekarakteriseerd door scanning-elektronenmicroscopie (SEM), röntgenenergie-dispersieve analyse (EDS) en atoomkrachtmicroscopie (AFM). Alkalische fosfatase (ALP) kleuring, Western blotting, qRT-PCR en immunofluorescentie kleuring werden uitgevoerd om het osteogene potentieel, het niveau van F-actine en de expressie van MKL1 en YAP/TAZ te onderzoeken. Onze resultaten toonden aan dat de binnendiameter en ruwheid van TiO2 nanobuisjes namen toe met de toename van de anodische oxidatiespanning van 30 naar 70 V, terwijl hun hoogte consistent 2 m was. Verder, hoe groter de buisdiameter, hoe sterker het vermogen van TiO2 nanobuisjes om osteogene differentiatie van MSC's te bevorderen. Het remmen van F-actine-polymerisatie door Cyto D remde osteogene differentiatie van MSC's evenals de expressie van eiwitten in focale adhesiecomplexen zoals vinculine (VCL) en focale adhesiekinase (FAK). Daarentegen verhoogde polymerisatie van F-actine na Jasp-behandeling de expressie van RhoA en transcriptiefactoren YAP/TAZ. Op basis van deze gegevens concludeerden we dat TiO2 nanobuisjes vergemakkelijkten de osteogene differentiatie van MSC's, en dit vermogen werd verbeterd met de toenemende diameter van de nanobuisjes binnen een bepaald bereik (30-70 V). F-actine bemiddelde dit proces via MKL1 en YAP/TAZ.
Inleiding
Titanium en titaniumlegeringen worden vanwege hun uitstekende biocompatibiliteit, corrosieweerstand en mechanische eigenschappen veel gebruikt in klinische toepassingen zoals totale gewrichtsvervangingen en tandheelkundige implantaten [1,2,3]. Er zijn echter nog veel uitdagingen die moeten worden opgelost, waaronder aseptische loslating en infectie [4, 5]. De afgelopen jaren zijn er een aantal onderzoeken uitgevoerd om de osseo-integratie en antibacteriële eigenschappen te verbeteren. Bijvoorbeeld de MoS2 /PDA-RGD-coating op titaniumimplantaten kan niet alleen de integratie van een titaniumimplantaat met het gastheerbot bevorderen, maar ook bacteriegroei met een hoge efficiëntie remmen [6]. Bovendien heeft de oppervlaktetopografie steeds meer aandacht getrokken, en topografische modificatie verschilt van chemische modificaties door alleen de structuur op micro- en nanoschaal te veranderen. De stimulatie van chemische signalen op cellen is onstabiel en cytotoxisch. Ter vergelijking:veilige en controleerbare fysieke signalen kunnen sommige bijwerkingen vermijden die worden veroorzaakt door chemische moleculen. Daarom kan topografische modificatie van het oppervlak van implantaten en de regulatie van osseo-integratie door topografische structuur een nieuwe manier bieden om het klinische probleem van slechte osseo-integratie na implantatie van prothesen op te lossen.
Op het gebied van botweefselengineering en botregeneratie wordt celmorfologie-interactie beschouwd als een veelbelovende managementstrategie voor nauwkeurige controle van de functie en differentiatie van zaadcellen. Tegelijkertijd heeft het bot zelf een elegante hiërarchie binnen het nanometer- en micronbereik [7]. Daarom kan oppervlaktemorfologie een vergelijkbare niche bieden, die de natuurlijke botstructuur kan nabootsen en osteogene differentiatie van mesenchymale stamcellen op het oppervlak van het gastheerbot en implantaat kan bevorderen. Oppervlaktemorfologieën kunnen uit veel verschillende structuren bestaan, waaronder nanobuisjes, nanodraden, nanoporiën, enzovoort. In het bijzonder hebben nanobuisarrays de afgelopen jaren veel belangstelling gewekt op veel gebieden vanwege hun unieke oppervlaktekenmerken, zoals een hoge oppervlakte-tot-volumeverhouding, biologische plasticiteit en hoge adsorptiecapaciteit. Een nieuwe studie toont bijvoorbeeld aan dat boornitride-nanobuisjes (BNNT) een gasgevoelig materiaal vormen dat kan worden gebruikt als gassensor om de werking van transformatoren te bewaken door de samenstelling en het gehalte van opgeloste gassen in olie te detecteren [8]. In de biogeneeskunde is oppervlaktetopografie ook in staat om cellulair gedrag te sturen, waaronder celmigratie, adhesie, proliferatie en differentiatie. De laatste onderzoeken laten zien dat topografie op nanoschaal mesenchymale stamcellen (MSC's) kan aansturen om te differentiëren tot osteoblasten om zo vroege osseo-integratie te versterken [9,10,11,12]. Er is zelfs gemeld dat gecombineerde oppervlaktemodificatie op micro- en nanoschaal ervoor kan zorgen dat MSC's differentiëren tot contractiele gladde spiercellen [13]. De moleculaire mechanismen van hoe oppervlaktetopografie het lot van cellen stuurt, moeten echter nog worden opgehelderd, wat belangrijk is voor de evaluatie van materiaalveiligheid en materiaalontwerp.
Filamenteuze (F)-actine, ook wel microfilament genoemd, is een van de drie belangrijkste componenten van het cytoskelet in eukaryote cellen. Het is samengesteld uit polymeren van bolvormig (G)-actine, gemodificeerd door tal van andere eiwitten. F-actine heeft structurele polariteit vanwege het feit dat alle subeenheden van het microfilament naar hetzelfde uiteinde wijzen. Het van weerhaken voorziene uiteinde is gericht op een ander aangrenzend monomeer, terwijl het puntige uiteinde een actine-subeenheid bezit met de ATP-bindingsplaats bloot. Dat wil zeggen, ATP is betrokken bij het proces van transformatie tussen G-actine en F-actine. Dit proces is in een dynamisch evenwicht, waarbij polymerisatie en depolymerisatie gelijktijdig plaatsvinden, ook bekend als loopbanden, vaak gezien bij lamellipodia en filopodia [14]. Daarom is het duidelijk dat actinedynamiek een belangrijke rol speelt in cellulaire functies zoals celmigratie, celdeling en behoud van celvorm. F-actine fungeert echter niet alleen als een fysieke structuur die mechanische belasting ondersteunt, maar neemt ook deel aan ander biologisch gedrag zoals signaaltransductie en genexpressie. Accumulerend bewijs toont aan dat F-actine fysieke signalen kan omzetten in chemische signalen door interactie met andere eiwitten [15,16,17,18]. Biomechanische en geometrische reconstructie bevorderen bijvoorbeeld de apoptose van tumorcellen door de superpositie van actinemonomeerpolymerisatie tot F-actine te voorkomen [15]. Ultrageluidpulsen verbeteren de osteogenese van menselijke mesenchymale stamcellen door de depolymerisatie van F-actine te remmen [16]. Onze eerdere studie toonde ook aan dat mechanische belasting de stabiliteit van F-actine verhoogt [17]. Omdat de holle structuur van nanobuisjes minder adhesieplaatsen voor cellen biedt, is herschikking van het cytoskelet onvermijdelijk om het biomechanische evenwicht te behouden. Daarom hebben we zeker reden om aan te nemen dat F-actine waarschijnlijk de door nanotopografie geïnduceerde celdifferentiatie medieert.
In deze studie hebben we TiO2 . gefabriceerd nanobuisjes, wijzigden hun topografie door anodische oxidatie en onderzochten hun vermogen om osteogene differentiatie van MSC's te bevorderen. Vervolgens hebben we onderzocht of F-actine een cruciale rol speelt bij mechanotransductie. Cytochalasine D (Cyto D), dat competitief bindt aan het van weerhaken voorziene uiteinde van F-actine om te voorkomen dat G-actine in het filament wordt opgenomen, werd gebruikt om de polymerisatie van F-actine te remmen, en jasplakinolide (Jasp) werd gebruikt om de stabilisatie van de actine montage. Verder wilden we ook ophelderen hoe F-actine functioneert bij het veranderen van fysieke signalen in biochemische signalen. Op basis van de resultaten van onze vorige studie veronderstelden we dat de MAPK-route mogelijk bij dit proces betrokken is [17]. Transcriptiefactoren zoals Ja-geassocieerd eiwit (YAP) / transcriptionele co-activator met PDZ-bindend motief (TAZ) en MKL1, die worden beschouwd als mechanosensoren en mechanotransducers, waren ook de onderwerpen van onze studie om te screenen hoe F-actine de stamcel beïnvloedt lot, omdat sommige onderzoeken op andere gebieden suggereerden dat ze gerelateerd waren aan F-actine [19,20,21]. In het algemeen hopen we de rol van F-actine in het proces van stamceldifferentiatie geïnduceerd door nanobuisjes te verduidelijken, om zo het materiaalontwerp en de bioveiligheidsbeoordeling van implantaten die zijn gemodificeerd door nanobuisjes te sturen.
Materialen en methoden
Vervaardiging van TiO2 Nanobuisjes
Plakken van puur titanium (99,9% zuiverheid, 2 mm dikte; Shengshida, Hebei, China), gebruikt als het substraat, werden gepolijst met siliciumcarbide schuurpapier van nr. 400 en 1500 grit. Monsters werden vervolgens achtereenvolgens gewassen met aceton, watervrije alcohol en gedeïoniseerd water in een ultrasone reinigingsmachine en tenslotte 3 uur bij kamertemperatuur gedroogd. Om de nanotopografie te fabriceren, werden de voorbehandelde monsters gefixeerd als de anode, terwijl een platinastuk als tegenkathode werd gebruikt in een waterige elektrolytoplossing van 0,15 M NH4 F en 90% glycol gedurende 1 uur. De anodisatiespanning was een constante spanning van 30, 40, 50, 60 of 70 V. Na anodische oxidatie werd elk monster 30 minuten gespoeld met gedeïoniseerd water en 15 minuten gewassen met watervrije alcohol in een ultrasone reinigingsmachine. Ten slotte werden alle monsters gedurende 1 uur in een autoclaaf bij 120 ° C gesteriliseerd en vóór gebruik bevochtigd met kweekmedium.
Het reactiemechanisme van de fabricage van nanobuisjes is niet duidelijk en de huidige gangbare theorie is de veldverbeterde oplossingstheorie. De vorming van nanobuisarrays is het resultaat van dynamisch evenwicht onder invloed van veldoxidatie, veldoplossing en chemische oplossing (figuur 1b). Het anodisatieproces kan als volgt worden beschreven:in de eerste stap wordt een oxidebarrièrelaag gevormd op het elektrolyt-metaalgrensvlak:
$$ {\mathrm{Ti}}^{4+}+2{\mathrm{H}}_2\mathrm{O}\to {\mathrm{Ti}\mathrm{O}}_2+4{\mathrm{ H}}^{+} $$ (1)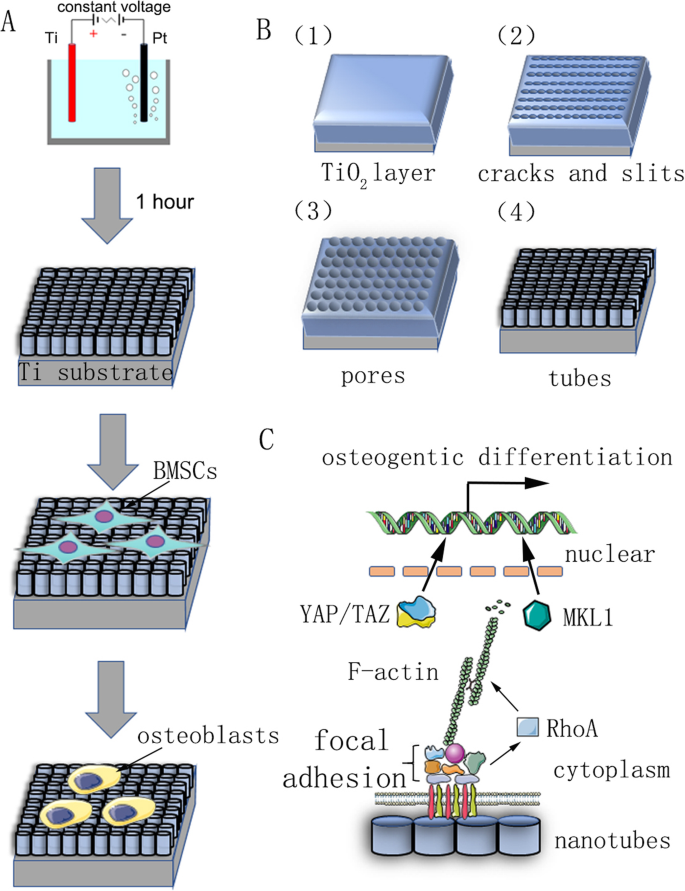
Een samenvattend overzicht van onze studie. een Stroomschema met anodische oxidatie en inductie van celdifferentiatie. b Het voorbehandelde titaniumsubstraat werd als anode gefixeerd in een waterige elektrolytoplossing met 0,15 M NH4 F en 90% glycol bij constante spanning gedurende 1 uur. Er werd voorspeld dat zelf-geassembleerde nanobuisjes uniform zouden worden gevormd. Het reactiemechanisme wordt beschreven in materialen en methoden. c Schematisch diagram van het mechanisme van osteogene differentiatie van stamcellen geïnduceerd door nanobuisjes
Dan verschijnen er barsten en smalle spleten aan het oppervlak als gevolg van veldversterkt oplossen van de oxidelaag. Verspreiding van F − ionen in deze scheuren en spleten verhoogt de oplossnelheid. Scheuren worden groter en raken verbonden met aangrenzende scheuren. Ten slotte bereiken de vormings- en oplossnelheid van de titaniumoxidelaag een dynamisch evenwicht en groeien de nanobuisjes niet meer:
$$ {\mathrm{TiO}}_2+6{\mathrm{F}}^{-}+4{\mathrm{H}}^{+}\to {\left[{\mathrm{TiF}}_6 \right]}^{2-}+2{\mathrm{H}}_2\mathrm{O} $$ (2)Oppervlaktekarakterisering
Monsters vervaardigd met verschillende spanningen (30, 40, 50, 60 of 70 V) werden 15 minuten gespoeld met ethanol en gedeïoniseerd water en vervolgens gedroogd bij kamertemperatuur. Scanning-elektronenmicroscopie (SEM450, FEI Nova Nano SEM; Thermo Fisher Scientific, Waltham, MA, VS) werd gebruikt om de oppervlaktestructuur te karakteriseren en om de binnendiameter en hoogte van de nanobuisjes te meten na het coaten van monsters met een dunne laag goud. Ondertussen werd X-ray energy dispersive analysis (EDS) uitgevoerd om de elementaire samenstelling van nanobuisjes te analyseren. Atoomkrachtmicroscopie (AFM, NanoManVS, Bruker Nano Surfaces, Bruker MicroCT, Kontich, België) werd gebruikt om de oppervlaktemorfologie en oppervlakteruwheid van de monsters te onderzoeken. Uit elk monster werden drie verschillende gebieden geselecteerd en de metingen werden drie keer herhaald.
Celcultuur
Vier weken oude mannelijke Sprague-Dawley (SD) ratten werden gekocht bij het proefdiercentrum van het Shanghai Ninth People's Hospital (Shanghai, China). Mesenchymale stamcellen van rattenbeenmerg (BMSC's) werden aseptisch geïsoleerd uit dijbenen en tibia's. BMSC's werden gezuiverd en verder geëxpandeerd in α-minimaal essentieel medium (α-MEM; Hyclone, Logan, UT, VS) dat 10% (v/v) foetaal runderserum (FBS) bevat (Gibco/Life Technologies, Carlsbad, CA, VS). ), 100 mg/ml streptomycine (Gibco) en 100 E/mL penicilline (Gibco), en geïncubeerd bij 37 °C in een bevochtigde atmosfeer bestaande uit 95% lucht en 5% CO2 . Het kweekmedium werd elke 2 dagen vernieuwd en de cellen werden getrypsiniseerd en bij 80% confluentie in subcultuur gebracht. Alle cellen die in deze studie werden gebruikt, bevonden zich tussen passage 3 en 5. Osteogeen inductiemedium bestond uit groeimedium aangevuld met 100 nM dexamethason, 10 mM β-glycerofosfaat en 50 mM ascorbinezuur (Sigma-Aldrich, St. Louis, MO, VS) .
Celproliferatietest
De TiO2 nanobuis-gemodificeerde titaniumplakjes werden in cirkelvormige vormen gesneden en in putjes van een celcultuurplaat met 24 putjes geplaatst. BMSC's tussen passages 3-5 werden gekweekt op TiO2 nanobuisjes met een dichtheid van 3 × 10 4 cellen/schijf in ofwel groeimedium of osteogeen medium. Na 2 dagen celcultuur werden cytochalasine D (Cyto D, Sigma-Aldrich) en jasplakinolide (Jasp, Sigma-Aldrich), gebruikt om de F-actine-polymerisatie te verstoren, elke dag gedurende 3 dagen aan het medium toegevoegd. De uiteindelijke concentratie en werktijd van Cyto D en Jasp waren respectievelijk 5 M, 1 uur en 2 M, 3 uur. Het kweekmedium werd vernieuwd na incubatie met de reagentia. De levensvatbaarheid en proliferatie van de cellen werden beoordeeld met behulp van de Cell Counting Kit-8 (CCK8)-assay (Dojindo, Kumamoto, Japan) 12 uur na behandeling met Cyto D of Jasp. De cellen werden 2 uur geïncubeerd met 10% (v/v) CCK8-oplossing in een celincubator bij 37 °C onder 5% CO2 . Vervolgens brachten we 100 L van het reactiemengsel over in putjes van een plaat met 96 putjes en de absorptie (OD) van het formazan-kleurstofproduct in de kweek werd gemeten bij 450 nm met behulp van een Multiscan UV-zichtbare spectrofotometer (Safire2; TECAN, Mannedorf, Zwitserland ). Daarnaast hebben we ook geschatte celaantallen geteld met behulp van een geautomatiseerde celteller (AMQAX1000, Life Technologies). Vóór het tellen van de cellen werden BMSC's enzymatisch losgemaakt van TiO2 nanobuisjes en gekleurd met Trypan-blauw (Sigma–Aldrich).
Alkalische fosfatasekleuring en ALP-activiteitsanalyse
BMSC's werden gezaaid op vijf verschillende TiO2 nanobuis-gemodificeerde titaniumschijfjes (30, 40, 50, 60 of 70 V) met een dichtheid van 3 × 10 4 per putje en gekweekt in osteogeen medium. Reagentia werden toegevoegd zoals hierboven beschreven. Na 7 dagen incubatie werden BMSC's gekweekt op TiO2 nanobuisjes werden drie keer gewassen met PBS, gefixeerd met 4% paraformaldehyde en geïncubeerd in alkalische fosfatase (ALP) werkoplossing van een ALP-kit volgens de instructies van de fabrikant (Hongqiao, Shanghai, China). De resultaten werden waargenomen onder een stereomicroscoop na wassen met PBS.
Voor ALP-activiteitsanalyse werden cellen eerst gelyseerd met RIPA-buffer zonder protease- en fosfataseremmers, en vervolgens werden de gecentrifugeerde lysaten getest met behulp van een ALP Assay Kit (Beyotime Institute of Biotechnology, Jiangsu, China) volgens het verstrekte protocol. De activiteit werd uiteindelijk genormaliseerd naar de eiwitconcentratie van het overeenkomstige lysaat.
Immunocytochemie
Na 3 dagen Cyto D- en Jasp-behandeling werden BMSC's gedurende 20 minuten bij kamertemperatuur gefixeerd met 4% paraformaldehyde en vervolgens driemaal gewassen met PBS. De cellen werden 30 min gepermeabiliseerd met 0,3% Triton-X 100, driemaal gewassen met PBS en gekleurd met Rhodamine-geconjugeerd phalloidin gedurende 1 uur bij kamertemperatuur in het donker. Vervolgens werden de cellen gespoeld met PBS en gedurende 10 minuten bij kamertemperatuur tegengekleurd met DAPI (Beyotime Institute of Biotechnology). Na drie extra wasbeurten met PBS werden de monsters op een glasplaatje gefixeerd en met confocale microscopie waargenomen.
Western Blotting
Om eiwitexpressie te evalueren, werden BMSC's gekweekt op TiO2 nanobuisjes werden geoogst met trypsine (Gibco). De cellen werden driemaal gewassen met PBS en gelyseerd met RIPA-buffer aangevuld met een cocktail van protease en fosfataseremmer gedurende 30 minuten op ijs. Het lysaat werd verzameld door centrifugatie bij 12.000 xg gedurende 15 minuten bij 4 ° C. De concentratie van totale eiwitten in het supernatant werd gemeten met behulp van een bicinchoninezuur (BCA) eiwittestkit (Beyotime) volgens de instructies van de fabrikant. Laadbuffer werd zoals hierboven aan eiwitmonsters toegevoegd, die vervolgens gedurende 15 minuten bij 95 ° C werden gekookt. Voor Western-blotanalyse werd 10 L van het eiwitpreparaat op een 12,5% SDS-PAGE-gel (EpiZyme Inc., Cambridge, MA, VS) geladen en gedurende 1 uur onderworpen aan elektroforese bij 120 V en vervolgens elektrisch overgebracht op een polyvinylideendifluoride (PVDF) membraan bij 250 mA gedurende 2 uur. De membranen werden vervolgens geblokkeerd met 5-10% magere melkpoeder in TBST gedurende 1 uur op een shaker bij kamertemperatuur en overnacht geïncubeerd met primair antilichaam verdund in verdunningsbuffer (Beyotime) bij 4 ° C. Vervolgens werd fluorescerend geconjugeerd secundair antilichaam verdund in verdunningsbuffer aan de membranen toegevoegd na driemaal wassen met TBST gedurende 5 minuten, en de membranen werden vervolgens gedurende 1 uur bij kamertemperatuur in het donker geïncubeerd. De eiwitbanden werden gedetecteerd door een tweekleurig infrarood fluorescentiebeeldvormingssysteem (Odyssey, LiCor Biosciences, Lincoln, NE, VS). In het bijzonder, als de banden van het interne referentie-eiwit verenigd waren, werd het membraan gestript en opnieuw onderzocht met een ander primair antilichaam, gevolgd door hetzelfde proces. We gebruikten GAPDH-antilichaam als ons interne referentie-eiwit om eiwitexpressie te normaliseren, en de andere primaire antilichamen die in deze studie werden gebruikt, waren anti-vinculine (1:1000 verdunning, Abcam, Cambridge, MA, VS), anti-FAK (1:1000 verdunning) , Celsignaleringstechnologie, Danvers, MA, VS), anti-Runx2 (1:1000 verdunning, celsignaleringstechnologie), anti-RhoA (1:1000 verdunning, celsignaleringstechnologie), anti-F-actine (1:500 verdunning) , Abcam), anti-Osx (1:500 verdunning, Abcam) en anti-pYAP (1:1000 verdunning, celsignaleringstechnologie). Secundaire antilichamen waren geiten-anti-muis IgG H&L (IRDye® 680RD, 1:5000 verdunning, Abcam) en geiten-anti-konijn IgG H&L (IRDye® 680RD, 1:5000 verdunning, Abcam).
Kwantitatieve realtime PCR
Kwantitatieve realtime PCR werd op dag 7 uitgevoerd om de genexpressie van runt-gerelateerde transcriptiefactor 2 (Runx2), Osterix (Osx), Alp, osteocalcine (OCN), RhoA, YAP, TAZ, vinculine (VCL), focale adhesiekinase (FAK) en megakaryoblastische leukemie 1 (MKL1) in cellen gekweekt in osteogeen medium op TiO2 nanobuisjes. Totaal RNA werd uit de cellen geëxtraheerd met behulp van een Total RNA Kit (R6812-01HP, Omega Bio-Tek Inc., Norcross, GA, VS). De concentratie en zuiverheid van RNA-monsters werden bepaald door optische dichtheid bij een golflengte van 260 en alleen monsters met zowel A260/280-verhoudingen als A260/230-verhoudingen hoger dan 1,8 werden geanalyseerd. RNA-monsters werden omgekeerd in cDNA getranscribeerd met behulp van een qScript cDNA-synthesekit (Takara, Shiga, Japan) volgens de instructies van de fabrikant. Kwantitatieve realtime PCR werd uitgevoerd met SYBR® Premix Ex Taq™ (Takara) met behulp van een QuantStudio 6 Flex realtime PCR-systeem (Life Technologies). GAPDH, een huishoudgen, werd als interne referentie gebruikt. Gegevens werden geanalyseerd met behulp van de vergelijking Ct (2 -ΔΔCt ) methode en uitgedrukt als vouwveranderingen vergeleken met de controle. De sequenties van de gebruikte primers staan vermeld in Tabel 1.
Statistische analyse
Alle gegevens zijn representatief voor ten minste drie onafhankelijke experimenten met monsters in drievoud, tenzij anders aangegeven. Gegevens worden uitgedrukt als het gemiddelde ± standaarddeviatie (SD). Verschillen tussen groepen werden geëvalueerd door eenrichtingsanalyse van variantie gevolgd door de Student-Newman-Keuls post-hoctest of Student's t test. P waarden < 0,05 werden als statistisch significant beschouwd.
Resultaten
Oppervlaktekarakterisering
Om de nanotopografie te fabriceren, TiO2 nanobuisjes werden gevormd op een puur titaniumsubstraat door anodische oxidatieapparatuur te gebruiken bij verschillende constante spanningen (30, 40, 50, 60 en 70 V) gedurende 1 uur (Fig. 1). Een uniform verdeelde reeks zelf-geassembleerde nanobuisjes werd waargenomen met behulp van scanning-elektronenmicroscopie (SEM). Zij- en bovenaanzichten van de nanobuisjes worden getoond in Fig. 2a, b. De hoogte van de nanobuisjes in alle monsters in dit onderzoek was ongeveer 2 m, terwijl de binnendiameters van de nanobuisjes ongeveer 74 nm (30 V), 92 nm (40 V), 112 nm (50 V), 128 nm (60 V) en 148 nm (70 V) (Fig. 2c). Dit gaf aan dat de hoogte van de nanobuisjes gerelateerd was aan de tijd van anodische oxidatie en dat de binnendiameter gerelateerd was aan de spanning van anodische oxidatie. Röntgenenergie-dispersieve analyse (EDS) werd vervolgens uitgevoerd om de elementaire samenstelling van de nanobuisjes te analyseren. Hieruit bleek dat de nanobuisjes uit slechts twee elementen bestaan, O en Ti (Fig. 2d, e). Atoomkrachtmicroscopie (AFM) werd gebruikt om de nanotubulaire structuren te detecteren en de rekenkundig gemiddelde afwijking van het profiel van nanobuisjes te meten als de oppervlakteruwheid (Ra) (Fig. 2f). De gegevens toonden aan dat de oppervlakteruwheid van nanobuisjes toenam naarmate de diameter (d.w.z. de spanning van anodische oxidatie) toenam (Fig. 2g).

Oppervlaktekarakterisering van nanobuisjes. een Zijaanzicht van nanobuisjes. Schaalbalken:1 m. b Bovenaanzicht van nanobuisjes. Schaalbalken:500 nm. c De binnendiameter van de nanobuisjes bij vijf verschillende constante spanningen (30, 40, 50, 60, 70 V). d De samenstelling van het EDS-chemische element van nanobuisstructuren. e De elementverhouding van Ti en O. f Atoomkrachtmicroscopie (AFM) beelden van nanobuisstructuren. g De gemiddelde oppervlakteruwheid (Ra) van de nanobuisjes
TiO2 Door nanobuisjes geïnduceerde osteogene differentiatie van MSC's
Na 7 dagen osteo-inductie werd eerst ALP-kleuring uitgevoerd om osteogene differentiatie van MSC's te evalueren. De kleurresultaten toonden aan dat MSC's gekweekt op TiO2 nanobuisjes hadden een hogere ALP-activiteit dan cellen gekweekt op een glad titaniumsubstraat (controlegroep) (figuur 3a). Statistische analyse van het kleuringsgebied toonde aan dat het vermogen van nanobuisjes om osteogene differentiatie te induceren significant was verbeterd in vergelijking met de controlegroep. Ondertussen hebben we een trend waargenomen dat binnen het diameterbereik van dit experiment, hoe groter de diameter van de TiO2 nanobuisjes, hoe sterker het vermogen om osteogene differentiatie te induceren (figuur 3b). Dus de 70 V-groep werd gebruikt in volgende experimenten om de resultaten beter weer te geven. Vervolgens analyseerden we osteogene genexpressie op dag 3 en 7. MSC's gekweekt op TiO2 nanobuisjes gedurende 3 dagen en 7 dagen vertoonden beide een significante bevordering van de expressie van osteogene genen (RUNX2, ALP, OCN en OSX) in vergelijking met de controlegroep (Fig. 3d-g). Western-blot-resultaten bevestigden dat de eiwitexpressie van RUNX2 en OSX ook toenam na 7 dagen osteo-inductie (figuur 3c). Interessant genoeg vonden we dat F-actine werd opgereguleerd in de TiO2 nanobuisjes groep. Daarom was het duidelijk dat TiO2 nanobuisjes stuurden MSC's naar osteoblastdifferentiatie, wat gerelateerd was aan de diameter van de nanobuisjes. Onze resultaten suggereerden ook de betrokkenheid van F-actine bij dit proces.

TiO2 nanobuisjes verbeterden osteogene genexpressie van BMSC's. een ALP-kleuring van het gladde titaniumsubstraat en vijf verschillende nanobuissubstraten. De cellen werden gedurende 7 dagen met osteogeen medium geïnduceerd. b Statistische analyse van het kleurgebied werd uitgevoerd met ImageJ. c Osteogenese-geassocieerde eiwitten (RUNX2 en OSX) en F-actine in MSC's werden geanalyseerd door Western-blotting op dag 7. De mRNA-expressie van RUNX2 (d ), ALP (e ), OCN (f ), en OSX (g ) op dag 3 en 7, geanalyseerd met qRT-PCR. NT de nanobuisgroep. Gegevens vertegenwoordigen het gemiddelde ± SD van drie monsters. *P <0,05, **P <0,01 en ***P <0,001
F-actine-gemedieerde osteoblastdifferentiatie van MSC's op TiO2 Nanobuisjes
Om verder te onderzoeken of F-actine betrokken was bij door nanotopografie geïnduceerde celdifferentiatie, gebruikten we twee reagentia, jasplakinolide (Jasp) en cytochalasine D (Cyto D), om de polymerisatie van F-actine respectievelijk op een positieve en negatieve manier te reguleren. De confocale microfoto's van Rhodamine-Phalloidin-kleuring toonden aan dat F-actine in de Cyto D-behandelingsgroep bijna gedepolymeriseerd was en vezelachtige structuren zelden werden gezien, terwijl Jasp F-actine stabiliseerde en polymeriseerde, geverifieerd door duidelijkere en helderdere bundelachtige structuren dan waargenomen in de controlegroep (Fig. 5a). Bovendien bevestigde Western-blot-analyse ook dat de eiwitexpressie van F-actine werd beïnvloed, wat aantoonde dat Cyto D en Jasp beide werkten zoals verwacht (Fig. 5b). Celproliferatietest toonde aan dat Cyto D de celproliferatie significant remde, terwijl Jasp de celgroei bevorderde (Fig. 4a). De resultaten van het celgetal waren consistent met deze bevinding (Fig. 4b).

F-actine-assemblage reguleerde de expressie van osteogene genen in BMSC's. een , b Celproliferatie na behandeling met Cyto D en Jasp werd bepaald met behulp van de CCK-8-assay of geautomatiseerde celteller op dag 1, 2 en 3. c , d ALP-kleuring en ALP-activiteitsanalyse werden uitgevoerd om de expressie van ALP in MSC's na medicamenteuze behandeling gedurende 3 dagen te evalueren. Het kleuringsgebied werd geanalyseerd met ImageJ. c , e –u Western blotting en qRT-PCR werden gebruikt om de veranderingen van osteogenese-gerelateerde markers in de NT+ Cyto D-groep en de NT+ Jasp-groep te vergelijken met die in de controlegroep (zonder medicamenteuze behandeling). NT de nanobuisjesgroep. Gegevens vertegenwoordigen het gemiddelde ± SD van drie monsters. *P <0,05, **P <0,01 en ***P <0,001
Vervolgens evalueerden we het vermogen van MSC's om te differentiëren tot osteoblasten om te onderzoeken of F-actine dit proces medieerde. We hebben voor het eerst ALP gedetecteerd als een vroege marker van osteogenese. In vergelijking met de controlegroep verminderde Cyto D-behandeling de expressie van ALP en zijn activiteit, terwijl die in de Jasp-behandelingsgroep werd opgereguleerd (Fig. 4c, d). In overeenstemming met dit resultaat resulteerde Jasp-behandeling in een verhoging van de eiwitniveaus van RUNX2 en OSX, terwijl Cyto D het tegenovergestelde effect had (Fig. 4c). In overeenstemming hiermee vertoonden de mRNA-expressieniveaus van osteo-specifieke genen, waaronder RUNX2, ALP, OCN en OSX, dezelfde trend na medicamenteuze behandeling (Fig. 4e-h). Bovenal gaven deze gegevens aan dat F-actine een belangrijke rol speelde in het proces van osteogene differentiatie van MSC's geïnduceerd door TiO2 nanobuisjes. Het bevorderen van F-actine-depolymerisatie remde de door nanotopografie geïnduceerde osteoblastdifferentiatie, terwijl de stabilisatie en polymerisatie van F-actine de osteoblastdifferentiatie verbeterde.
F-actine gereguleerde osteoblastdifferentiatie van MSC's op TiO2 Nanobuisjes via MKL1 en YAP/TAZ
Om het onderliggende mechanisme van F-actine bij de regulatie van het lot van MSC's te ontleden, hebben we eiwitten / moleculen onderzocht die direct interageren met F-actine of de polymerisatie van F-actine beïnvloeden. Eerst probeerden we te identificeren hoe de nanotopografie de balans tussen F-actine en G-actine beïnvloedde. TiO2 nanobuisjes als een fysiek signaal verschillen van membraanpermeabele chemische signalen en moeten dus enkele componenten van het celmembraan gebruiken om stimuli in de cellen over te brengen. Steeds meer bewijs geeft aan dat het focale adhesiecomplex, inclusief integrine, talin, focale adhesiekinase (FAK), vinculine (VCL), tensine en andere eiwitten, functioneert als een signaaldrager, die cellen informeert over de toestand van de extracellulaire matrix (ECM). ) en beïnvloedt zo hun biologisch gedrag [22, 23]. Wat nog belangrijker is, F-actine bindt aan integrines via dergelijke focale adhesiecomplexen en vormt daardoor mechanische verbindingen tussen intracellulaire actinebundels en de ECM [24]. Daarom analyseerden we vervolgens de expressie van componenten van het focale adhesiecomplex. De resultaten toonden aan dat de eiwit- en mRNA-expressie van VCL en FAK consistent waren met de verandering van F-actine, wat aangeeft dat het focale adhesiecomplex betrokken was bij het proces van osteogene differentiatie van MSC's geïnduceerd door TiO2 nanobuisjes (figuren 5b en 6a, b). Daarnaast vonden we ook dat RhoA, een klein GTPase-eiwit in de Rho-familie van GTPasen, werd opgereguleerd in de Jasp-behandelingsgroep en geremd door Cyto D (figuren 5b en 6a, b). RhoA is een belangrijk stroomopwaarts signaaltransductiemolecuul in de MAPK-route en zou kunnen worden gereguleerd door FAK [25, 26]. De primaire functie van RhoA is het bevorderen van de polymerisatie en stabiliteit van spanningsvezels (F-actine) en de assemblage van het focale adhesiecomplex [27]. Alles bij elkaar suggereerden deze gegevens dat TiO2 nanobuisjes kunnen de polymerisatie van F-actine beïnvloeden via het focale adhesiecomplex en RhoA.

Immunofluorescentiekleuring onthulde het niveau van F-actine door kleuring met rhodamine-geconjugeerd phalloidin (a ). The protein expression of FAK and VCL contained in the focal adhesion complex, RhoA and phosphorylated YAP were investigated by Western blotting (b ). NT the nanotubes group

Effect of F-actin assembly on FAK (a ), vinculin (VCL) (b ), RhoA (c ), MKL1 (d ), YAP (e ), and TAZ (f ) gene expression in MSCs. NT the nanotubes group. Data represent the mean ± SD of three samples. *P <0.05, **P <0.01, and ***P <0,001
But how does F-actin regulate cell fate? Most studies have demonstrated that F-actin is involved in cell migration, cell division, endocytosis, and especially tumor cell invasion [28,29,30]. Few studies have suggested that F-actin could also regulate cell differentiation, let alone its specific molecular mechanism [31, 32]. Consequently, we searched for articles that mentioned the F-actin changes and found that YAP/TAZ, two closely related transcriptional co-activators in the Hippo signaling pathway, which shuttle between the cytoplasm and the nucleus, may serve as mechanotransducers in regulating MSC differentiation [33,34,35]. In addition, we also found that MKL1, a key regulator of smooth muscle cell differentiation, which interacts with the transcription factor serum response factor, could bind to G-actin and also circulate between the cytoplasm and the nucleus [21, 36]. Our results ultimately identified the involvement of YAP/TAZ and MKL1 in nanotube-induced osteoblast differentiation mediated by F-actin (Figs. 5b and 6d–f). Interestingly, the protein expression of phosphorylated YAP showed the opposite trend, indicating that not only was the expression of YAP changed, but the phosphorylation of YAP was also changed by Cyto D and Jasp (Fig. 5b). This result was consistent with the report that the phosphorylation of YAP/TAZ could be sequestrated in the cytoplasm [35].
In summary, our results preliminarily demonstrated that F-actin regulated osteoblast differentiation of MSCs on TiO2 nanotubes through MKL1 and YAP/TAZ (Fig. 7).

Schematic representation of F-actin assembly induced by nanotubes, and the putative role of MKL1 and YAP/TAZ in acting as the downstream mediators of F-actin signaling to regulate gene expression
Discussie
Titanium and titanium alloys are the most widely used metal materials in orthopedic clinical implants due to the good properties of titanium [3]. However, aseptic loosening is still an urgent problem to be solved and improved, and the key is likely to lie in improving the integration of the implant and the host bone. Previous studies have shown that surface coating and modification or immobilization of biofunctional molecules will be beneficial to osseointegration [37]. Recently, the surface topography of implants has attracted the attention of many researchers thanks to studies into the cell response to physical cues [9,10,11, 13, 38]. In this study, we demonstrated the ability of nanotubes to promote osteogenic differentiation of MSCs, and this ability was enhanced with increasing inner diameter of the nanotubes (30–70 V). This will help guide the diameter of the nanotubes on the surface of the implants.
As a topographical structure, nanotubes first change the physical properties of the material, such as adsorption capacity and electrical and thermal conductivity. These physical properties determine their application in the industrial field. For example, most high-voltage power transformers need to be filled with insulating material, which is usually transformer oil or insulating gas. When the insulation of a transformer fails due to overheating and partial discharge, a serious discharge accident will occur. Therefore, finding an effective method that accurately detects the concentration and types of dissolved gases or insulating gas decomposition components in a transformer is necessary to monitor the operating state of the transformer [8, 39,40,41]. The traditional approach is to look for materials with good gas adsorption in transition elements, which are rich in d electrons, such as Pd(1 1 1) [39]. Nowadays, nanotubes are widely studied for their good gas adsorption properties. He et al. found that CuO-BNNT was suitable for the adsorption of C2 H2 , because of its stronger adsorption on C2 H2 [8]. Meanwhile, TiO2 itself can be a gas-sensing material. Gui et al. found that Co-doped TiO2 further enhanced gas adsorption capacity and exhibited a superior adsorption ability and conductivity change toward C2 H4 molecules [40]. Consistent with this study, Mn-doped graphene also exhibited enhanced conductivity and superior capability of C2 H2 and CO detection than pristine graphene [41]. The above research indicates that the TiO2 nanotubes prepared in our experiment have a potential application in the field of monitoring the operative state of a transformer. However, the adsorption capacity and electrical conductivity of the nanotubes to gases need to be further studied, especially whether these properties are enhanced after doping with transition elements (e.g., Mn).
In addition to changing the physical properties of a surface, nanoscale morphology also affects the biological behavior of the cells attached to it. Cells first adhere to the surface of the material and then migrate, proliferate, and differentiate. Compared to a flat surface, the hollow structure of the nanotubes provides fewer adhesion sites for cells. Therefore, in order for the cells to adhere to the nanotube surface steadily and maintain the biomechanical balance within the cell, the focal adhesion complex begins to assemble and mature, and F-actin becomes strong and stable.
F-actin, a linear polymer microfilament consisting of G-actin monomers, is one of the three major components of the cytoskeleton. As a mechanical-loading structure, F-actin is generally believed to be involved in cell division, cell migration, endocytosis, and tumor cell invasion [28,29,30], but some recent studies showed that it can also affect cell differentiation [31,32,33, 36]. For example, actin cytoskeletal depolymerization by simvastatin induces chondrocyte differentiation [31], and actin depolymerization enhances adipogenic differentiation in human stromal stem cells [32]. Our results also revealed that, compared with the control group, MSCs cultured on nanotubes had higher F-actin levels and a more obvious fibrous structure. Meanwhile, promotion of F-actin polymerization by Jasp enhanced osteogenic differentiation, while the depolymerization of F-actin inhibited osteogenic differentiation, suggesting that F-actin mediates TiO2 nanotube-induced osteoblastic differentiation of MSCs.
F-actin can be regulated by Rho GTPases, members of the Ras superfamily [23, 42], and Rho can induce actin reorganization through at least two effectors, ROCK and Dia. ROCK is activated by binding to Rho-GTP and then myosin light chain (MLC), the substrate of ROCK, plays an important role in F-actin assembly. ROCK inhibits the activity of MLC phosphatase, leading to an increase in MLC phosphorylation, which stimulates the ATPase activity of myosin II and promotes the assembly of F-actin. In addition, ROCK also targets LIM kinase (LIMK). Phosphorylated LIMK inactivates cofilin by phosphorylation, which can disassemble F-actin in its active state. Another effector is Dia, a member of the formin-homology (FH) family of proteins which contains two FH domains. These domains contain multiple proline-rich motifs which bind to the G-actin-binding protein, profilin. This interaction contributes to actin polymerization and F-actin organization [42]. We detected one of the Rho GTPases, RhoA, and found that the expression of RhoA was consistent with the level of F-actin. However, we were unable to clearly describe how the nanotubes regulate the expression of RhoA, because there are many other regulators, including integrin signaling, other adhesion receptors, G protein-coupled receptors (GPCRs), soluble factors such as LPA, receptor tyrosine kinase signaling, and so on [43].
Knowing that F-actin can be regulated by RhoA, we next asked what role focal adhesion played in this process, because focal adhesion complexes, containing integrins, talin, vinculin, paxillin, and focal adhesion kinase (FAK), are formed and mature when cells attach to the surface of nanotubes. Integrins are transmembrane heterodimers that couple the ECM to the other focal adhesion proteins so as to facilitate cell attachment. They not only act simply as hooks but also transmit to the cell critical signals about the nature of its surroundings, which along with other signals such as EGFR, prompt the cell to make decisions about its biological behaviors. These signals are further transmitted to F-actin, which is directly connected to the focal adhesion complexes. On the one hand, the nanoscale morphology causes focal adhesion complex assembly and maturation. On the other hand, kinases such as FAK and Src kinase family members will recruit molecules such as CRK to self-regulate the assembly and maturation of focal adhesion complexes [44,45,46]. Our results demonstrated that the formation and maturation of focal adhesion complexes were impaired by F-actin depolymerization, suggesting that there was a feedback from focal adhesion complexes to actin assembly in line with published reports.
However, it should not be ignored that these proteins contained in focal adhesion complexes have the function of signal transduction [47]. That is to say, nanotubes may directly regulate gene expression through signal cascades, and F-actin may just participate in or be affected by this process. For instance, the dual kinase complex of FAK and Src can regulate Rho GTPases such as RhoA. This shows that nanotubes can regulate RhoA through integrins and the FAK/Src complex. In addition Src, a non-receptor tyrosine kinase protein, can activate Ras (small GTPase) by phosphorylating FAK at tyrosine residue 925 [47, 48]. Then, Ras activates numerous biochemical pathways, including the well-studied MAPK pathway and the PI3K/AKT/mTOR pathway. In the MAPK pathway, Ras activates c-Raf, followed by mitogen-activated protein kinase kinase (MAP2K) and then MAPK1/2, also known as extracellular signal-regulated kinase (ERK). ERK in turn activates transcription factors such as serum response factor (SRF) and c-Myc that are involved in regulating growth and differentiation [49]. What is more, Runx2, a key transcription factor in osteogenic differentiation, can also be regulated by ERK [50], and our previous study confirmed that mechanical strain promoted osteogenic differentiation of BMSCs through the FAK-Erk1/2-Runx2 pathway [17]. Therefore, we cannot rule out that ERK plays a role in nanotube-induced osteogenic differentiation and further study is still needed.
So what exactly is the role of F-actin in inducing differentiation of nanotubes, because its change can affect cell differentiation? One possibility is that the change of F-actin assembly can inversely regulate the level of FAK so as to induce osteogenic differentiation through the FAK-Erk1/2-Runx2 pathway as described above, because in our results, focal adhesion complexes and actin polymerization showed the same trends of change, indicating that they act as a whole in response to the extracellular environment. However, some other possibilities also exist, and a number of articles have shown that MKL1 and YAP/TAZ act downstream of the actin dynamic balance [20, 51,52,53,54]. Both of them shuttle between the cytoplasm and the nucleus, and may help to transduce signals from the cytoskeleton to the nucleus.
MKL1, also termed myocardin-related transcription factor A, is sensitive to changes in G-actin levels. When cytoplasmic G-actin levels increase, monomeric G-actin binds to MKL1 and prevents it from binding to SRF and activating transcription. SRF target genes include actins such as smooth muscle actin (SMA) as well as other actin-binding proteins, including immediate early genes like c-fos and egr1. Recent studies have demonstrated that changing SRF activity could regulate adipogenesis by activating the adipogenesis transcription factor peroxisome proliferator-activated receptor γ (PPARγ), and also regulate bone formation via IGF-1 and Runx2 signaling [55, 56].
YAP and TAZ are two transcriptional coactivators in the Hippo signaling pathway, identified as an important regulatory pathway that restricts cell proliferation, thereby controlling organ size and morphogenesis [20]. Large tumor suppressor genes 1 and 2 (LATS1/2) phosphorylate them, thereby creating a binding site for 14-3-3 proteins, the binding of which prevents their nuclear import [53, 54]. As a consequence, phosphorylated forms of YAP/TAZ are sequestered in the cytoplasm, preventing the expression of genes like Ctgf and Areg. In addition, some studies have shown that YAP/TAZ can interact with T-box 5 (TBX5), RUNX2, and p73 to regulate gene expression [57,58,59]. Further, cell adhesion to cell matrix proteins has been shown to trigger YAP nuclear localization through an integrin/FAK/Src axis. In our study, the results suggested that this pathway was possibly involved in nanotube-induced differentiation. Further study into the downstream mediators of the integrin/FAK/Src axis should be carried out to clarify the specific mechanism.
On the other hand, more and more studies illustrate that F-actin interacts with Hippo signaling, and somehow inhibits the phosphorylation of YAP [54, 60], which is consistent with our experimental results that promoting F-actin polymerization reduces the expression of phosphorylated YAP. We hypothesize that ATP involved in the process of the transformation between G-actin and F-actin may also play an important role in the phosphorylation of YAP, which is yet to be studied.
After understanding the above possible molecular mechanisms, we can try to explain some of the experimental phenomena found in this study. Our results revealed that the larger the diameter of the nanotubes, the stronger the ability of the nanotubes to promote osteogenic differentiation. This is consistent with previous research [61, 62]. The reason for this phenomenon is that the larger the diameter of the nanotubes, the less adhesion sites they can provide to the cells, and the greater the assembly and maturity of focal adhesion complexes. Along with these, stress fibers made of F-actin will have greater strength and stability. These structures enhance the signaling that promotes osteogenic differentiation. Predictably, however, this effect is significantly reduced when the nanotubes become too large in diameter, making it difficult for the cells to adhere to the surface [12]. Similarly, when the height of the nanotubes is inconsistent, the differences in height can result in a change of adhesion site and rearrangement of the cytoskeleton, which will further affect cell differentiation. Intriguingly, even flat surface materials without nanotube modification can induce changes in cell differentiation. A number of studies have demonstrated that focal adhesion formation and stress fiber organization are regulated by substrate stiffness [63,64,65], and YAP/TAZ also plays an important role in this process. Therefore, it is obvious that the integrins–FAs (focal adhesions)–F-actin axis plays a role in the transduction of physical signals into intracellular chemical signals.
In summary, our results demonstrated that F-actin regulates osteoblastic differentiation of mesenchymal stem cells on TiO2 nanotubes through MKL1 and YAP/TAZ, whose target genes partly explained the proliferation and differentiation of MSCs. We know that there is no single change in the signal network and any change is regulated by numerous molecules and proteins. One type of biological behavior must be the result of the regulation of a series of signaling pathways. Nanotubes induce cell differentiation by triggering a complex network of signals, including integrins, proteins contained in focal adhesion complexes, FAK, Src, Rho GTPase, the MAPK pathway, the Hippo pathway, and other reported signaling pathways. At least as important, there are many signal cycles in the signal network and a downstream signal can regulate the upstream signal via feedback. In this study, we found that vinculin and FAK can be regulated backwards by F-actin assembly, increasing the uncertainty of molecular function. Therefore, more details of the molecular mechanism await further study.
Conclusies
Our results showed that TiO2 nanotubes promoted the osteogenic differentiation of MSCs, and this ability was enhanced with the increasing diameter of nanotubes within a certain range (30–70 V). F-actin mediated nanotube-induced cell differentiation through MKL1 and YAP/TAZ, providing a novel insight into the study of cell differentiation.
Beschikbaarheid van gegevens en materialen
De datasets die tijdens het huidige onderzoek zijn gebruikt en geanalyseerd, zijn op redelijk verzoek verkrijgbaar bij de corresponderende auteur.
Afkortingen
- MSCs:
-
Mesenchymal stem cells
- SEM:
-
Scanning elektronenmicroscopie
- EDS:
-
X-ray energy dispersive analysis
- AFM:
-
Atoomkrachtmicroscopie
- ALP:
-
Alkaline phosphatase
- Cyto D:
-
Cytochalasin D
- Jasp:
-
Jasplakinolide
- VCL:
-
Vinculin
- FAK:
-
Focal adhesion kinase
- BCA:
-
Bicinchoninic acid
- PVDF:
-
Polyvinylidenedifluoride
- Runx2:
-
Runt-related transcription factor 2
- Osx:
-
Osterix
- OCN:
-
Osteocalcin
- YAP:
-
Yes-associated protein
- MKL1:
-
Megakaryoblastic leukemia 1
- FBS:
-
Foetaal runderserum
- ECM:
-
Extracellular matrix
- MLC:
-
Myosin light chain
- LIMK:
-
LIM kinase
- FH:
-
Formin-homology
- GPCR:
-
G protein-coupled receptors
- MAP2K:
-
Mitogen-activated protein kinase kinase
- ERK:
-
Extracellular signal-regulated kinase
- SRF:
-
Serum response factor
- SMA:
-
Smooth muscle actin
- PPARγ:
-
Peroxisome proliferator-activated receptor γ
- LATS1/2:
-
Large tumor suppressor gene 1 and 2
- TBX5:
-
T-box 5
Nanomaterialen
- TITANIUMDIOXIDE - TiO2 - Prijs, markt en analyse
- Op weg naar TiO2-nanovloeistoffen:deel 1:voorbereiding en eigenschappen
- Nanotechnieken inactiveren kankerstamcellen
- Op weg naar TiO2-nanovloeistoffen - Deel 2:Toepassingen en uitdagingen
- Effect van gouden nanodeeltjesdistributie in TiO2 op de optische en elektrische kenmerken van kleurstofgevoelige zonnecellen
- Synthese van ZnO-nanokristallen en toepassing in omgekeerde polymeerzonnecellen
- De optimale titaniumvoorloper voor het vervaardigen van een TiO2-compacte laag voor perovskietzonnecellen
- Invloed van elastische stijfheid en oppervlakteadhesie op het stuiteren van nanodeeltjes
- Invloed van Ag-nanodeeltjes met verschillende afmetingen en concentraties ingebed in een TiO2-compacte laag op de conversie-efficiëntie van perovskiet-zonnecellen
- Een nieuw up-conversiemateriaal van Ho3+-Yb3+-Mg2+ tri-gedoteerde TiO2 en zijn toepassingen op perovskiet-zonnecellen
- Invloed van koolstofnanobuisjes en zijn derivaten op tumorcellen in vitro en biochemische parameters, cellulaire bloedsamenstelling in vivo