


Beoordeling van door gouden nanodeeltjes geremde cytochroom P450 3A4-activiteit en moleculaire mechanismen die ten grondslag liggen aan de cellulaire toxiciteit ervan in humane hepatocellulaire carcinoomcellijn C3A
Abstract
Interacties van de 40 en 80 nm gouden nanodeeltjes (AuNP) gefunctionaliseerd met kationisch vertakt polyethyleenimine (BPEI), anionisch liponzuur (LA) of neutrale polyethyleenglycol (PEG) met humaan hepatocellulair carcinoom (HCC) cellijn C3A zijn onderzocht in de afwezigheid en aanwezigheid van humaan plasma-eiwit corona (PC). Alle kale (geen PC) AuNP behalve 80 nm LA-AuNP waren cytotoxisch voor C3A, maar PC verzwakte hun cytotoxiciteit. Tijdsafhankelijke cellulaire opname van AuNP nam toe naast 40 nm BPEI-AuNP, maar PC onderdrukte hun opname naast 80 nm PEG-AuNP. Bifasische reacties van oxidatieve / nitrosatieve stress door BPEI-AuNP kwamen voor in C3A-cellen, terwijl PEG-AuNP een krachtige antioxidant was. Alle kale AuNP remden de cytochroom P450 (CYP) 3A4-activiteit, ongeacht de grootte en oppervlaktelading, maar PC recupereerde zijn activiteit naast PEG-AuNP. De 40 nm PEG-AuNP-gemoduleerde genexpressie was voornamelijk betrokken bij mitochondriale vetzuur β-oxidatie en in mindere mate hepatische efflux/opname transporters. Deze onderzoeken dragen bij aan een beter begrip van AuNP-interactie met belangrijke biologische processen en hun onderliggende moleculaire mechanismen in HCC, die mogelijk verder betrokken zijn bij de ontwikkeling van een effectiever therapeutisch doelwit bij HCC-behandeling.
Achtergrond
Hepatocellulair carcinoom (HCC) is een van de meest voorkomende vormen van kanker wereldwijd en de snelst groeiende doodsoorzaak door kanker in de VS [1, 2]. Aangezien HCC in een vergevorderd stadium is gediagnosticeerd, omvatten de curatieve HCC-behandelingen levertransplantatie of chirurgische resectie bij de vroege tumorontwikkeling en chemo- en radiotherapie voor een vergevorderde tumortoestand. HCC ontwikkelt vaak een hoge resistentie tegen conventionele antineoplastische middelen, een niet-selectieve cytotoxische molecule die systemische bijwerkingen zou kunnen veroorzaken. De recente ontwikkelingen in gentherapie, d.w.z. op RNA-interferentie (RNAi) gebaseerde gentherapie, zijn gebruikt in de huidige HCC-behandeling [3, 4]. De werkzaamheid van RNAi vereist dat de vector wordt afgeleverd in het inwendige van de doelcel [5]. De vectoren voor een succesvolle genafgifte zijn virale en niet-virale vectoren. Virussen bieden een grotere efficiëntie van genafgifte, maar niet-virale vectoren hebben de voorkeur vanwege veiligheidsproblemen met de virale vectoren. Nanodeeltjes (NP) als niet-virale vectoren voor gerichte genafgifte of geneesmiddelafgiftesysteem hebben veel aandacht gekregen voor het verbeteren van de therapeutische efficiëntie en het verlagen van de toxiciteit op systemische en/of cellulaire niveaus bij HCC-behandeling [4, 6]. Het wordt dus erg belangrijk om het moleculaire mechanisme en de biologische route te identificeren die ten grondslag liggen aan cellulaire verstoring en toxiciteit van NP in doelcellen en weefsels. Recente in vitro-onderzoeken hebben aangetoond dat genexpressieprofilering in combinatie met cellulaire en biochemische reacties heeft geleid tot een directe beoordeling van cellulaire verstoring en potentiële NP-toxiciteit [7,8,9,10].
De gouden nanodeeltjes (AuNP) zijn gebruikt als leveringsvehikel voor doelspecifieke levering van gen-uitschakelingsgroepen, alleen of in combinatie met andere geneesmiddelen vanwege hun unieke fysisch-chemische eigenschappen en oppervlaktechemie [11, 12]. AuNP-interactie met bloedplasma-eiwitten vormt eiwitcorona, dat op zijn beurt de NP-oppervlaktechemie verandert en de daaropvolgende biologische reacties zoals de cellulaire opname en potentiële toxiciteit beïnvloedt [13, 14]. Cellulaire opnames van AuNP in verschillende menselijke kankercellijnen en primaire cellen werden kritisch beïnvloed door de vorming van corona-eiwitten, ongeacht de grootte en oppervlaktelading [7,8,9, 14,15,16,17].
De grootte- en oppervlakteladingsafhankelijke oxidatieve stress werd ook waargenomen in menselijke borstkankercellijn, MDA-MB-231, hepatocellulair carcinoom HepG2 en menselijke leukemie HL-60-cellen als reactie op AuNP, die geassocieerd waren met NP-cytotoxiciteit [18] , 19]. AuNP-geïnduceerde cytotoxiciteit trad op in verschillende menselijke kankercellijnen en primaire menselijke cellen op een celtype-specifieke manier [7,8,9, 20, 21].
Cytochroom P450 (CYP) -enzymen spelen een belangrijke rol bij de bioactivering of inactivering van talrijke cytotoxische geneesmiddelen en de gevoeligheid van de gastheer voor de carcinogeniteit van geneesmiddelen tegen kanker [22]. AuNP beïnvloedde de katalytische activiteit van CYP-enzymen op cellulair en moleculair niveau in vivo en in vitro [7, 23,24,25]. AuNP heeft aanzienlijk de differentiële genexpressie aangetoond die voornamelijk betrokken is bij oxidatieve stressmarkers in menselijke longfibroblastcellijn MRC-5, en mitochondriale disfunctie in menselijke navelstrengadercellen (HUVEC) en menselijke hepatocyten, wat correleert met een toename van de productie van lipideperoxide en een hoge cytotoxiciteit [8, 9, 26]. Hoewel deze kennis wederkerig suggereert dat AuNP apoptotische of necrotische celdood veroorzaakt in verschillende celtypes en cellulaire en biochemische functies verandert in combinatie met differentiële genexpressie in stressresponsroutes en toxiciteit, zijn de specifieke routes waardoor AuNP hun toxische effecten uitoefent in de cel of biologische systeem blijft onbekend.
Hierin onderzocht deze studie de effecten van eiwitcorona, grootte en oppervlaktelading op AuNP-interactie met menselijke HCC-cel C3A. In de eerste plaats werd de tijdafhankelijke cellulaire opname van de 40 en 80 nm AuNP gefunctionaliseerd met kationisch BPEI, anionisch liponzuur (LA) of neutraal polyethyleenglycol (PEG) in C3A-cellen bepaald met en zonder corona van menselijk plasma-eiwit (PC). Ten tweede werden AuNP-geïnduceerde cytotoxiciteit en productie van reactieve zuurstofsoorten (ROS) / reactieve stikstofsoorten (RNS) gevolgd samen met hun remmende effecten op CYP3A4-activiteit. Ten slotte werd het met AuNP-toxiciteit geassocieerde moleculaire werkingsmechanisme gekarakteriseerd met behulp van de Human Molecular Toxicology Pathway Finder en de Human Drug Transporters RT 2 Profiler™ PCR-array.
Methoden
Synthese van gouden nanodeeltjes
De 40 en 80 nm kationische BPEI, anionische LA en neutrale PEG Biopure ™ AuNP werd op maat gesynthetiseerd uit nanoComposix (San Diego, CA). Deeltjesgrootte, polydispersiteitsindex (PDI) en zeta (z)-potentiaal en spectrale eigenschappen werden gekarakteriseerd met dynamische lichtverstrooiing (DLS), transmissie-elektronenmicroscopie (TEM) en UV-Vis-spectroscopie. AuNP werden gesynthetiseerd door de reductie van waterstoftetrachloorauraat (III) hydraat in waterige kaliumcarbonaatoplossing gevolgd door het verouderingsproces en tangentiële stroomfiltratie (TFF). AuNP-oppervlak werd gefunctionaliseerd met LA of PEG door toevoeging van dihydroliponzuur (0,2:1, w) /w ) of thiol-methoxy-getermineerde PEG (Laysan Bio Inc., Arab, AL) (0,5:1, w /w ), respectievelijk gevolgd door wassen met TFF en steriele filtratie. BPEI-gefunctionaliseerde oppervlakken van AuNP werden gesynthetiseerd via EDC-chemie door het carbonzuur van LA te koppelen aan vrije aminen van BPEI, gevolgd door wassen met TFF en daaropvolgende centrifugatie voor een verwijdering van ongebonden BPEI.
Eiwit Corona-voorbereiding
Samengevoegd menselijk bloedplasma (n = 5) werden verkregen van de Biological Specialty Corp. (Colmar, PA). AuNP werden geïncubeerd in humaan plasma (55%, v /v ) bij een constante snelheid van 250 tpm bij 37 °C gedurende 1 uur zoals gerapporteerd [7, 8]. De ongebonden en zwak geassocieerde eiwitten werden verwijderd door herhaald wassen met fosfaatgebufferde zoutoplossing (PBS) bij 20.000 xg gedurende 20 min bij 20 °C. De uiteindelijke met menselijke plasma-eiwit corona (PC) gecoate AuNP werd gedispergeerd in PBS en vervolgens verdund in celkweekmedium voor verdere fysisch-chemische karakterisering of dosering. Het gedetailleerde protocol wordt gegeven in het aanvullende bestand 1.
Fysisch-chemische karakterisering van AuNP
Hydrodynamische diameters (D H ), PDI en z-potentiaal van de 40 en 80 nm kale (geen pc) AuNP gefunctionaliseerd met BPEI, LA en PEG in gedeïoniseerd (DI) water werden geanalyseerd bij 25 ° C bij 0 uur met behulp van de Zetasizer Nano ZS (Malvern Instruments, Worcestershire, VK); voor PC-gecoate AuNP in PBS bij 25 ° C na 0 uur; en voor alle kale en PC AuNP in volledig celkweekmedium bij 37 ° C bij 0 uur en 24 uur. Compleet celkweekmedium bevatte Eagle's minimum essential medium (EMEM) aangevuld met 10% FBS (ATCC ® , Manassas, Virginia). Een monster werd 5 keer gemeten met 11 subruns van elk 10 s. Bovendien werden optische absorptiespectra gemeten met behulp van de Synergy H1 hybride multi-mode microplaatlezer (BioTek Instruments Inc., Winooski, VT) bij kamertemperatuur om 0 uur.
Transmissie-elektronenmicroscoop
De AuNP-morfologie werd gekarakteriseerd met behulp van TEM. Alle kale en PC AuNP-oplossing (5 L) werd op koperen roosters van 200 mesh geplaatst, gevolgd door drogen aan de lucht bij kamertemperatuur. De monsters werden bekeken op een Tecnai G2 Spirit BioTWIN met een Oxford-detector (FEI Company, Hillsboro, OR) bij een versnellingsspanning van 120 kV. De GATAN-microscopiesuite (GATAN Inc., Pleasanton, CA) heeft AuNP-diameters gemeten.
Meting van celcultuur en levensvatbaarheid
Humaan hepatocellulair carcinoom C3A-cellen (ATCC ® CRL-10741™) zijn gekocht bij ATCC ® (Manassas, VA), gekweekt in volledige EMEM (ATCC ® , Manassas, VA) aangevuld met 10% FBS, en geëxpandeerd tot ongeveer 80% confluentie in T75-kolf met mediumverversingen om de 4 dagen. Na 0,25% (w /v ) trypsine-0,53 mM ethyleendiaminetetraazijnzuur (EDTA) digestie, cellen werden uitgeplaat in platen met 96 putjes bij 8 × 10 4 cellen per putje en geïncubeerd bij 37 °C in een bevochtigde atmosfeer van 95% lucht en 5% CO2. Na 48 uur incubatie werden cellen gedoseerd met AuNP in de afwezigheid en aanwezigheid van PC. De C3A-cellen tussen passage 9 en 12 werden gebruikt voor de dosering.
De levensvatbaarheid van C3A werd bepaald met behulp van de alamarBlue ® levensvatbaarheidstest (Thermo Sci., Waltham, MA) zoals beschreven [7, 27]. Cellen in de platen met 96 putjes werden behandeld met de 40 en 80 nm BPEI-, LA- en PEG-AuNP met en zonder pc variërend van 0 tot 250 μg/cm 2 . Na 24 uur, 10% alamarBlue ® reagens in volledige EMEM (v /v ) werd aan de celcultuur toegevoegd en gedurende 3 uur bij 37 ° C geïncubeerd. Het volledige EMEM diende als dispergeermiddel. De interacties van AuNP met het actieve ingrediënt van alamarBlue ® reagens, resazurin of een gereduceerd product, resorufine werden gemeten als controles. AuNP en resazurin (geen cellen) of onderhoudsmedium (geen cellen) dienden als achtergrondcontroles. Fluorescentie, evenredig met de levensvatbaarheid van de cellen, werd genormaliseerd naar controles en uitgedrukt als een percentage ten opzichte van de controlecelgroep.
Meting van celopname met inductief gekoppelde plasma-massaspectrometrie
Cellen werden gezaaid bij 8 × 10 4 cellen per putje van platen met 96 putjes en gedoseerd met een niet-toxische concentratie van 1,56 μg/cm 2 van alle kale en PC AuNP voor 0,5, 1, 3, 6, 12 en 24 uur. De etsstap werd opgenomen om celmembraangebonden AuNP en zijn niet-specifieke binding aan de putjes te verwijderen, zoals eerder gerapporteerd [28]. De celoogst werd gedroogd en verteerd in koningswater en de intracellulaire Au-concentratie werd gekwantificeerd met behulp van de NexION ™ 350X inductief gekoppelde plasma-massaspectrometrie (ICP-MS) (PerkinElmer, Waltham, MA). Cellulaire opname van AuNP werd berekend zoals eerder gerapporteerd en uitgedrukt als het aantal AuNP per cel [29]. Het gedetailleerde protocol wordt gegeven in het aanvullende bestand 1.
Oxidatieve/nitrosatieve stressmetingen
Cellen werden gezaaid bij 8 × 10 4 cellen per putje van platen met 96 putjes en gedoseerd met de 40 nm kale BPEI- en PEG-AuNP tot 125 μg/cm 2 voor 1, 3 en 24 uur. Directe meting van zuurstof/nitrosatieve stress werd getest met totaal reactieve zuurstofspecies (ROS)/superoxide (SO) testkit (Enzo Life Sciences, Farmingdale, NY) zoals eerder beschreven [30]. Fluorescentie, evenredig met de toename van ROS/reactieve stikstofspecies (RNS) (Ex488/Em520 nm) of SO (Ex550/Em610 nm) werd gemeten met de microplaatlezer. Het gedetailleerde protocol wordt gegeven in het aanvullende bestand 1.
Cytochrome P450 3A4-activiteit
Nadelige effecten van de 40 en 80 nm kale en PC AuNP op CYP3A4-activiteit werden gekarakteriseerd met behulp van P450-Glo ™ -assays (Promega Corp., Madison, WI) zoals volledig beschreven [7]. C3A-cellen in platen met 96 putjes werden gedoseerd bij de mediane letale concentratie (LC50 ) waarden:127,3 μg/cm 2 van de 40 nm BPEI-AuNP, 205,5 μg/cm 2 van de 80 nm BPEI-AuNP, 192,5 μg/cm 2 van de 40 nm LA-AuNP en 129,5 μg/cm 2 van de 40 nm PEG-AuNP. Sinds LC50 waarden van de 80 nm LA- en PEG-AuNP werden niet bepaald, cellen werden behandeld met LC50 waarden van de 40 nm LA- en PEG-AuNP (192,5 μg/cm 2 en 129,5 μg/cm 2 respectievelijk). Na het einde van de incubatie van 24 uur werden de cellen gedurende 3 uur bij 37 ° C geïncubeerd met een CYP3A4-substraat (luciferine-IPA). Het luminescentiesignaal, evenredig met een enzymactiviteit, werd gemeten met een microplaatlezer en vervolgens genormaliseerd naar controles. Controles werden toegewezen om de interactie van AuNP met oudersubstraten of metabolieten en celvrije substraten te beoordelen. CYP-activiteit werd uitgedrukt als een percentage ten opzichte van de controlecelgroep.
Genexpressieprofilering
Omdat toxisch PEG-AuNP van 40 nm werd gebruikt bij de remming van CYP3A4-activiteit en antioxidantactiviteit in C3A-cellen die een hoge cellulaire opname vertonen, werd het geselecteerd om moleculaire werkingsmechanismen te karakteriseren die ten grondslag liggen aan de toxiciteit en differentiële cellulaire reacties. Cellen werden gezaaid bij 2.5 × 10 6 cellen per putje van platen met 6 putjes en gedoseerd met LC50 waarde van de 40 nm PEG-AuNP gedurende 24 uur bij 37 ° C. Aan het einde van de incubatie werden cellen onderworpen aan RNA-isolatie en vervolgens werd cDNA-synthese uitgevoerd met behulp van totaal RNA met een gemiddelde RNA-integriteitsgetallen (RIN)-waarde van 7,8 zoals eerder beschreven [7,8,9]. Het resulterende cDNA werd gemengd met RT 2 SYBR groene mastermix (Qiagen Inc., Valencia, CA) en vervolgens toegepast op de Human Molecular Toxicology Pathway Finder of Human Drug Transporters RT 2 Profiler™ PCR-arrays in Quantstudio™ 7 Flex (Applied BioSystem, Foster City, CA). Differentieel tot expressie gebrachte genen met de vouwverandering <− 2 en> 2 en a p <-0,05 vertegenwoordigde neerwaartse en opwaartse regulatie van het gen van belang. Om de RT 2 te valideren PCR-arraygegevens, een expressie van negen geselecteerde genen werd geëvalueerd met cDNA-synthese en daaropvolgende realtime PCR. Primersequenties zijn samengevat in Aanvullend bestand 1:Tabel S1. Alle PCR-reacties werden in drievoud uitgevoerd. Het gedetailleerde protocol van real-time PCR-condities en kwantificering wordt gegeven in het aanvullende bestand 1.
Statistische analyse
Mediane letale concentratie (LC50 ) waarden van AuNP in C3A-cellen werden geschat door een Hill-vergelijking met variabele helling aan te passen aan de waargenomen gegevens (de invoer van AuNP-concentratieniveaus en de bijbehorende levensvatbaarheid van de cellen) met behulp van GraphPad Prism 6 (La Jolla, CA) zoals beschreven [7]. Eenrichtingsanalyse van variantie (ANOVA) werd uitgevoerd met behulp van SAS 9.4 (SAS Institute, Cary, NC) om de effecten van AuNP op ROS / RNS-productie en cellulaire opname in C3A-cellen te beoordelen. Indien significant, werd de meervoudige vergelijking uitgevoerd met Tukey's eerlijke significante verschil (HSD) test op een p < 0.05.
Resultaten en discussie
Fysicochemische karakterisering van kale en menselijke plasma-pc AuNP
De effecten van NP-grootte, oppervlaktelading en PC-vorming in menselijk plasma rond AuNP op de hydrodynamische diameter (DH ), polydispersiteitsindex (PDI), z-potentiaal en een spectrale eigenschap, evenals de morfologie, zijn gekarakteriseerd met behulp van DLS-, TEM- en UV-Vis-spectroscopie (Fig. 1). In TEM-afbeeldingen was alle kale (geen pc) AuNP in DI-water monodispers met de strakke grootteverdeling en unieke UV-Vis-spectrumbereiken van 521-553 nm (figuur 1a). PC-formaties rond AuNP werden waargenomen met de veranderingen in grootteverdeling en roodverschuivingen van de absorptiespectra (figuur 1b). De D H en PDI-waarden van de 40 en 80 nm kale en PC AuNP in volledige EMEM waren compatibel tot 24 uur bij 37 °C, behalve voor de 40 en 80 nm PC BPEI-AuNP die een afname in PDI-waarden liet zien (respectievelijk 0,29 en 0,32). ) na 24 uur bij 37 °C vergeleken met die na 0 uur bij 37 °C (respectievelijk 0,62 en 1,0) (tabel 1). Z-potentiaalwaarden van alle kale en PC AuNP daalden relatief na 24 uur bij 37 ° C in vergelijking met die na 0 uur bij 37 ° C. Een aggregatie van de 40 en 80 nm PC BPEI-AuNP in PBS en complete EMEM werd waargenomen, wat correleerde met meerdere pieken in een grootteverdeling en veranderingen in DH en roodverschuivingen van absorptiespectra ten opzichte van kale BPEI-AuNP (Fig. 1 en aanvullend bestand 1:Figuur S1, Tabel 1). Deze resultaten werden ondersteund door de recente onderzoeken dat de 40 en 80 nm pc en het humaan serumalbumine corona-gecoate BPEI-AuNP werden geaggregeerd in PBS en verschillende celkweekmedia [7,8,9].
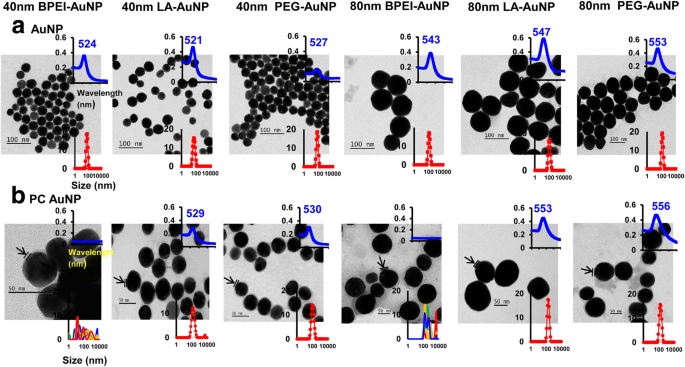
Transmissie-elektronenmicrofoto's van a AuNP in gedeïoniseerd water en b PC AuNP in PBS bij 0 uur bij 25 ° C, UV-Vis spectra golflengte (bovenste inzet), en de dynamische lichtverstrooiing distributie (onderste inzet). Pijlen geven pc-vorming aan. PC humaan plasma-eiwit corona, BPEI vertakt polyethyleenimine, LA liponzuur, PEG polyethyleenglycol
AuNP-cytotoxiciteit
AuNP-cytotoxiciteit werd gemeten met behulp van mediane letale concentratie (LC50 ) in C3A-cellen. De NP-oppervlaktelading-, deeltjesgrootte- en PC-vorming rond NP-afhankelijke LC50 analyses met AuNP werden getoond in Fig. 2. Alle 40 nm BPEI-, LA en PEG-AuNP en de 80 nm BPEI-AuNP waren cytotoxisch voor C3A-cellen met de overeenkomstige LC50 varieert van 127,3 tot 205,5 μg/cm 2 (Fig. 2a). De 80 nm kale PEG-AuNP vertoonde een levensvatbaarheid van 59% van de cellen bij de hoogste concentratie van 250 μg/cm 2 , terwijl de 80 nm LA-AuNP niet cytotoxisch was. PC verminderde AuNP-toxiciteit als functie van grootte en oppervlakteladingmodificatie, behalve de 80 nm BPEI-AuNP die 51% cellevensvatbaarheid vertoonde bij 250 μg/cm 2 om 24 uur (Fig. 2b). Recente onderzoeken hebben aangetoond dat de 40 nm kale BPEI-AuNP toxisch was voor primaire humane hepatocyten, HUVEC en humane renale proximale tubulaire cellen (HPTC) (LC50 varieert van 22,4–80,3 μg/cm 2 ) [7,8,9]. De PC-gecoate BPEI-AuNP was cytotoxisch voor humane hepatocyten, maar HSA-gecoate AuNP was niet cytotoxisch [7]. Deze resultaten suggereerden dat C3A-cellen resistenter zijn tegen AuNP-toxiciteit dan primaire menselijke cellen vanwege een hoge proliferatiesnelheid en metabolische activiteit van kankercellijn [31].
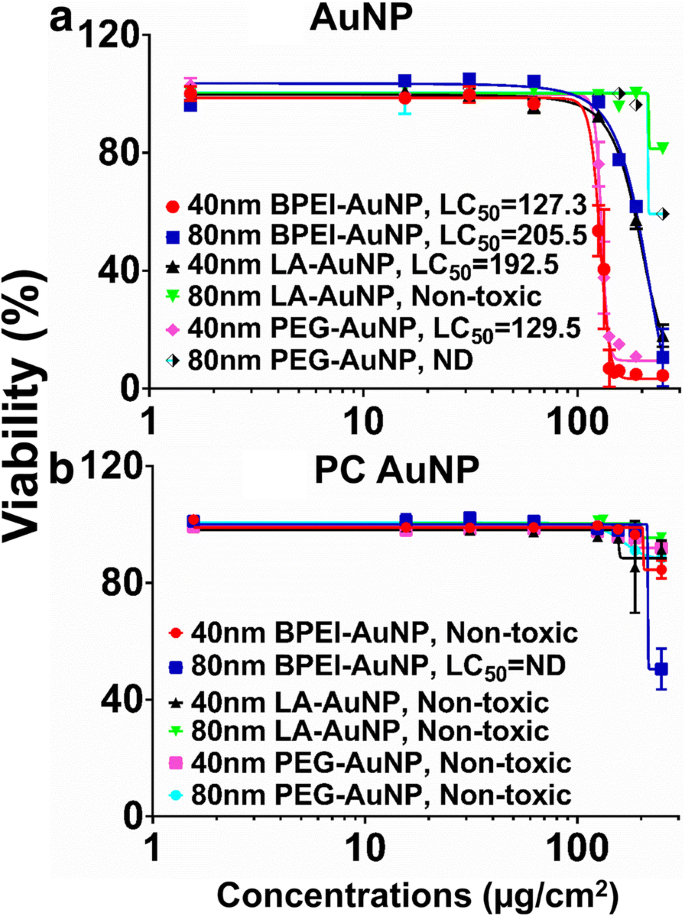
C3A levensvatbaarheid en LC50 waarden van de 40 en 80 nm a AuNP en b PC AuNP. Gegevens vertegenwoordigen gemiddelde ± S.D. (n = 3). PC humaan plasma-eiwit corona, ND niet bepaald, BPEI vertakt polyethyleenimine, LA liponzuur, PEG polyethyleenglycol, LC50 mediane dodelijke concentratie
Intracellulaire opname van AuNP
NP-grootte-, oppervlaktelading- en pc-afhankelijke cellulaire opname van alle kale en pc-AuNP werden bepaald op 1,56 μg/cm 2 tot 24 uur. ANOVA toonde significante veranderingen met grootte, pc en tijd (p < 0,0001), en interacties (pc × grootte, pc × tijd, grootte × tijd en pc × grootte × tijd) (p < 0,001) voor alle AuNP-opname behalve een onbeduidende interactie (PC × grootte) voor LA- en PEG-AuNP-opname (p = 0.2). Zoals getoond in Fig. 3a-f, werd een lineaire toename in cellulaire opname van de 40 en 80 nm kale en PC AuNP waargenomen naast de 40 nm kale en PC BPEI-AuNP die de hoogste cellulaire opname bereikte na 6 uur en daarna afnam ( Afb. 3a). Na 24 uur bevatte de 40 nm kationische BPEI-AuNP echter de hoogste opname gevolgd door neutrale 40 nm PEG-AuNP en vervolgens anionische 40 nm LA-AuNP, die geassocieerd was met de volgorde van C3A-celcytotoxiciteit van AuNP (Fig. 2a ). Dit resultaat is consistent met de eerdere studies dat kationisch poly (N-(2-aminoethyl)acrylamide)- en BPEI-AuNP de grootste cellulaire opname hadden vergeleken met die van anionisch poly(acrylzuur)- en LA-AuNP en neutraal poly( N-(2,3-dihydroxypropyl)acrylamide- en PEG-AuNP in humane colorectale adenocarcinoom Caco-2-cellen, HPTC en humane hepatocyten [9, 32]. Bovendien verzwakte het NP-PC-complex alle 40 en 80 nm BPEI- en LA-AuNP en de 40 nm PEG-AuNP in C3A-cellen maar versnelde de 80 nm PEG-AuNP-opname (Fig. 3f). Deze resultaten worden ondersteund door de recente onderzoeken dat PC de opname van AuNP in HUVEC, HEK en HPTC remde, ongeacht de grootte en oppervlaktelading [8, 9, 33]. Daarentegen versterkten PC- en HSA-corona's de 40 nm PEG-AuNP-opname in menselijke hepatocyten, maar die laatste induceerde de 80 nm PEG-AuNP-opname in HEK [7, 33] .
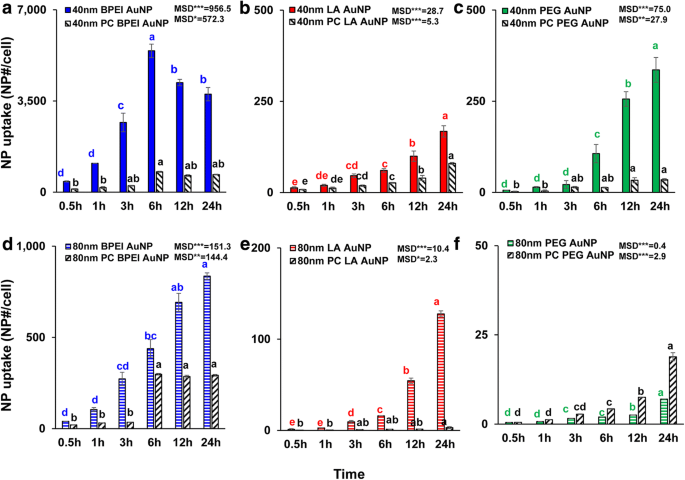
Tijdsafhankelijke mobiele opname van de 40 nm a BPEI-AuNP, b LA-AuNP en c PEG-AuNP en de 80 nm d BPEI-AuNP, e LA-AuNP en f PEG-AuNP bij afwezigheid en aanwezigheid van pc in C3A-cellen tot 24 uur. Gegevens vertegenwoordigen gemiddelde ± S.D. (n = 3). Letters waren significant verschillend volgens Tukey's HSD-test. BPEI vertakt polyethyleenimine, LA liponzuur, PEG polyethyleenglycol, PC humaan plasma-eiwit corona, MSD minimaal significant verschil. *p < 0,05; ** p < 0,005; ***p < 0.0001
Metingen van oxidatieve en nitrosatieve stress
Omdat de 40 nm kale BPEI- en PEG-AuNP de hogere cytotoxiciteit en cellulaire opname in C3A-cellen vertoonden in vergelijking met andere AuNP, werden ze geselecteerd om AuNP-geïnduceerde oxidatieve / nitrosatieve stress te onderzoeken. Beide AuNP moduleerden de ROS/RNS-generatie in C3A-cellen op tijd- en concentratieafhankelijke wijze (p < 0,0001) en door interactie (tijd × concentratie, p < 0,0001). Zoals weergegeven in figuur 4a, nam de ROS/RNS-generatie af bij de hogere concentraties van het 40 nm BPEI-AuNP (62,5 μg/cm 2 en 125 μg/cm 2 ) na 1 uur bij 37 ° C, maar nam toe tot 24 uur. Daarentegen onderdrukte de 40 nm PEG-AuNP de ROS/RNS-generatie aanzienlijk bij 39,1 μg/cm 2 verder met een vouwverandering < 0,5 tot 24 uur (Fig. 4b). Activering van celdood draagt vaak bij aan NP-toxiciteit en in de meeste gevallen is een toename van de ROS/RNS-productie, wat leidt tot oxidatieve stress, verantwoordelijk voor NP-toxiciteit [34]. De oppervlakteladingsafhankelijke ROS / RNS-productie werd waargenomen met de 40 nm kationische BPEI- en neutrale PEG-AuNP. De 40 nm BPEI-AuNP vertoonde een bifasisch patroon van ROS / RNS-generatie (antioxidant na 1 uur en pro-oxidant na 3 uur) bij hoge concentraties, wat geassocieerd was met zijn cytotoxiciteit in C3A-cellen (Fig. 2a). Dit resultaat is consistent met de eerdere onderzoeken dat de 40 en 80 nm BPEI-AuNP en de 20 nm citraat-AuNP-geïnduceerde ROS-generatie was geassocieerd met hun cytotoxiciteit in respectievelijk menselijke hepatocyten en HepG2-cellen, op tijd- en concentratieafhankelijke wijze [7, 35]. AuNP toonde oxidatieve stress-geïnduceerde cytotoxiciteit in humane promyelocytische leukemiecellen, HL-60 met een totale glutathionreductie, ongeacht de grootte [19]. Daarentegen diende de 40 nm PEG-AuNP als een antioxidant, wat suggereert dat oxidatieve/nitrosatieve stress mogelijk geen direct mechanisme is van de 40 nm PEG-AuNP-geïnduceerde cytotoxiciteit in C3A-cellen (Fig. 2b).
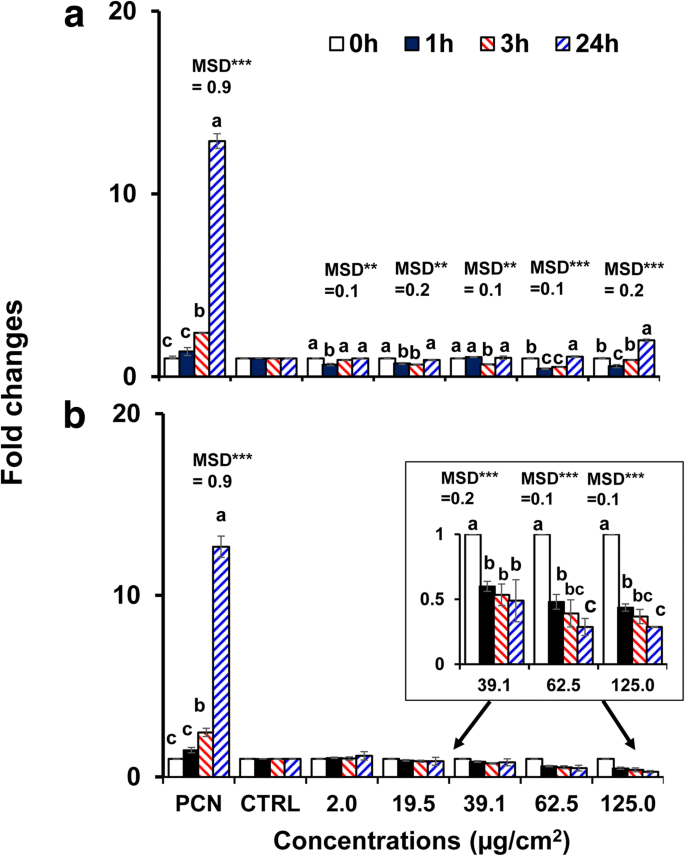
Tijd- en concentratieafhankelijke ROS/RNS-productie in C3A-cellen die zijn blootgesteld aan a de 40 nm BPEI-AuNP en b de 40 nm PEG-AuNP tot 24 uur. Gegevens vertegenwoordigen gemiddelde ± S.D. (n = 3). Letters waren significant verschillend volgens Tukey's HSD-test. BPEI vertakt polyethyleenimine, LA liponzuur, PEG polyethyleenglycol, CTRL controle, MSD een minimaal significant verschil, PCN pyocyanine (ROS-inductor). ** p < 0,005; ***p < 0.0001
CYP3A4-activiteitsmeting
Remmende effecten van de 40 en 80 nm kale en PC AuNP op CYP3A4-activiteit werden gekarakteriseerd. Zoals weergegeven in figuur 5, de 40 nm BPEI-, LA- en PEG-AuNP en de 80 nm BPEI-AuNP bij LC50 waarden remden de katalytische activiteit van CYP3A4 in C3A-cellen met de overeenkomstige activiteit van 20,1 tot 31,4% ten opzichte van de controles, ongeacht de grootte en oppervlaktelading. Niet-toxische concentraties van de 80 nm LA- en PEG-AuNP onderdrukten ook de activiteit (respectievelijk 31,4 en 26,6%). PC verbeterde echter aanzienlijk de 40 en 80 nm AuNP-geïnduceerde CYP3A4-remming naast de 40 en 80 nm PEG-AuNP die 63 en 46% activiteit vertoonde in vergelijking met de controles. Dit komt overeen met in vitro-onderzoeken met humaan leverweefsel en hepatocyten dat anionisch looizuur-AuNP en kationisch 40 en 80 nm BPEI-AuNP de katalytische activiteit van CYP3A4 aanzienlijk remden [7, 25]. Daarentegen kationische PEI-AuNP en neutrale polyvinylpyrrolidon-AuNP-geïnduceerde mRNA-expressie van CYP1A2, CYP2C9 en CYP3A4 in HepG2-cellen en CYP2B en CYP3A in respectievelijk rattenleverplakjes [36, 37]. Recent onderzoek meldt dat de 40 en 80 nm kale en PC BPEI-AuNP de CYP3A4-activiteit in menselijke hepatocyten substantieel onderdrukte via een conformationele verandering in eiwit of het blokkeren van de substraatpocket als een omkeerbare remming [7].
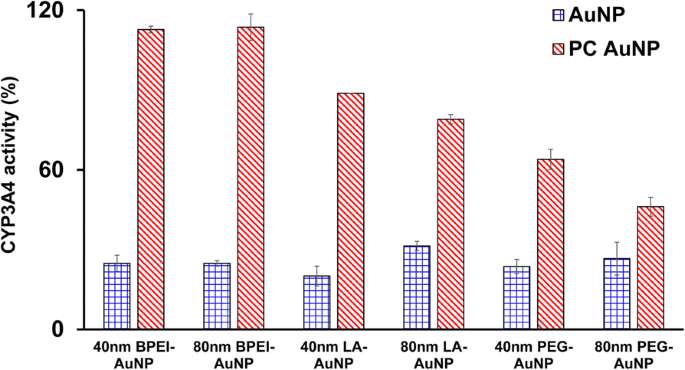
Een remmend effect van AuNP op CYP3A4-activiteit in C3A-cellen die gedurende 24 uur zijn blootgesteld aan de 40 en 80 nm BPEI-, LA- en PEG-AuNP in afwezigheid en aanwezigheid van pc. Waarden vertegenwoordigen gemiddelde ± S.D. (n = 3). BPEI vertakt polyethyleenimine, LA liponzuur, PEG polyethyleenglycol, PC humaan plasma-eiwit corona
Toxiciteitstrajectgerichte genexpressieprofilering van de 40 nm PEG-AuNP
Van de representatieve genen die 13 verschillende stress- en toxiciteitsroutes omvatten, werden in totaal 212 genen (↓186 en ↑26 genen) differentieel tot expressie gebracht op LC50 waarde van de 40 nm PEG-AuNP (afb. 6, aanvullend bestand 1:tabellen S2-S7). De 12,3% (26 genen, ↓26, ↑0 genen) van de totale genen (212 genen) waren voornamelijk betrokken bij mitochondriale vetzuur β-oxidatie; voor apoptose 11,3% (24 genen, A18, A6 genen); voor DNA-beschadiging en herstelroute 11,3% (24 genen, ↓18, ↑6-genen); en voor hitteschokrespons 11,3% (24 genen, ↓22, ↑2).
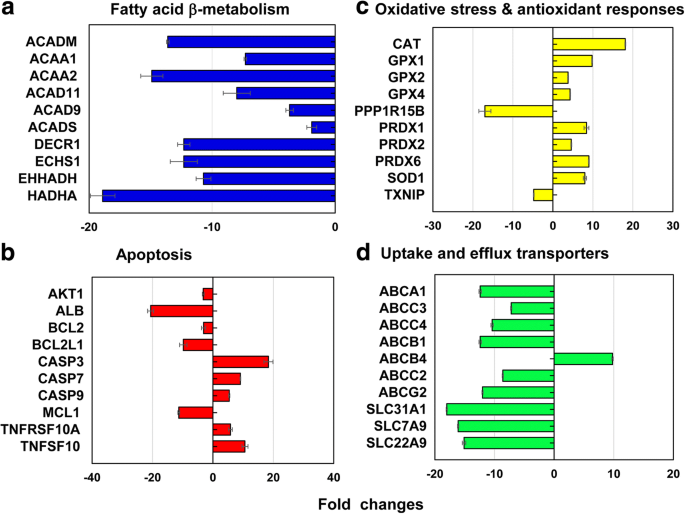
Representatieve genen betrokken bij a mitochondriaal vetzuur β-oxidatie, b apoptose, c oxidatieve stress en antioxidantreacties, en d leveropname en effluxtransporters bij LC50 waarde van 40 nm PEG-AuNP. Alle gegevens hadden een vouwverandering <− 2 en> 2 bij p < 0.05. Genontologie-analyse wordt vermeld in Aanvullend bestand 1:Tabellen S2-S7
In de mitochondriale vetzuur-β-oxidatieroute, genen die coderen voor drie verschillende enzymen die betrokken zijn bij de productie van acyl-CoA en het verminderen van equivalenten van NADH en FADH2 werden voornamelijk onderdrukt; ACAD11-, ACAD9-, ACADM- en ACADS-genen in Acyl-CoA-dehydrogenasen (2,0 tot 13,6-voudig); ACAA1 en ACAA2 in ketoacyl-CoA-thiolasen (7,3- tot 14,9-voudig); DECR1, ECHS1, EHHADH en HADHA (10,7- tot 18,9-voudig) in enoyl-CoA-hydratase (figuur 6a, aanvullend bestand 1:tabel S2). Mitochondriale vetzuur-β-oxidatie speelt een belangrijke rol bij de productie van acyl-CoA en het verminderen van equivalenten van NADH en FADH2 , dat is geassocieerd met vier hoofdenzymen (acyl-CoA-dehydrogenasen, enoyl-CoA-hydratasen, hydroxyacyl-CoA-dehydrogenasen en ketoacyl-CoA-thiolasen [38, 39]. Verder elektronendragers, NADH en FADH2 , zijn betrokken bij de tricarbonzuur (TCA) -cyclus en de mitochondriale ademhalingsketen, wat resulteert in ATP-productie. In de huidige studie induceerde de 40 nm PEG mitochondriale disfunctie, een verlies van ATP-onderhoud via een afname van de intracellulaire niveaus van ATP en FADH2 , waardoor de cytotoxiciteit ervan in C3-cellen wordt gedefinieerd (figuur 2a). Het vergelijkbare fenomeen werd gerapporteerd in menselijke hepatocyten, HUVEC en HPTC, blootgesteld aan het 40 nm BPEI-AuNP, wat aangeeft dat mitochondriale disfunctie een algemeen mechanisme van AuNP-toxiciteit kan zijn, ongeacht de oppervlaktelading en celtypes [7,8,9]. Een recente studie heeft gemeld dat mitochondriale disfunctie-relevante cytotoxiciteit werd waargenomen in onsterfelijk gemaakte prostaatkankerepitheelcellen en longkankerepitheelcellen als reactie op een remmer van STAT3-fosforylering, OPB-51602 [40].
In de apoptose-route werden de zes pro-apoptotische genen van CASP3, CASP7, CASP9, TNFRSF10A, TNFRSF10B en TNFSF10 opgereguleerd, terwijl de zes anti-apoptotische genen van AKT1, ALB, BCL2, BCL2L1, MCL1 en XIAP werden gedownreguleerd (Fig. 6b, Aanvullend bestand 1:Tabel S3), dat gecorreleerd was met dosisafhankelijke cytotoxiciteit in C3A-cellen (Fig. 2a). In DNA damage and repair check point, genes of the checkpoint kinases (CHEK1/2), the DNA excision repair genes (ERCC1/2/3), and the DNA ligase IV (LIG4) were upregulated but other excision repair genes (ERCC5/6, XRCC1/5), the checkpoint kinase (CDKN1A), and protein kinases (PRKDC) genes were downregulated (2- to 19-fold). These results suggested that the 40 nm PEG-AuNP-induced interference with cell cycle and DNA repair system may correlate with an induction of cell death in C3A cells (Fig. 2a, Additional file 1:Table S3). Genes encoding two different heat shock proteins (HSP) (A1A and A1B) were upregulated (10.2- to 14.2-fold) but HSP40 subfamily A, B, and C; HSP90 member 1; and HSP60 were downregulated (2- to 16-fold) (Additional file 1:Table S4).
In oxidative stress and antioxidant response, the 40 nm PEG-AuNP at LC50 value induced antioxidants genes and suppressed pro-oxidants, which was associated with a decrease in ROS/RNS generation being antioxidant itself (Fig. 4b). In antioxidant genes, glutathione peroxidase (GPX) 1, GPX2, GPX4, PRDX1, PRDX2, PRDX6, superoxide dismutase (SOD) 1, and CAT were induced (3.8- to 18.1-fold). In pro-oxidant genes, TXNIP and PPP1R15B were suppressed (4.8- and 17-fold, respectively) (Fig. 6c, Additional file 1:Table S5). This is consistent with a previous study that AuNP displayed oxidative stress-induced cytotoxicity in HepG2 and human hepatocytes, irrespective of size [7, 19].
In phase I metabolism, CYP3A4 and ESD genes were extensively suppressed (7-fold and 12-fold, respectively). Especially, inhibitory effect of 40 nm PEG-AuNP on CYP3A4 expression was correlated with a decrease in CYP3A4 activity (Fig. 5). Recent studies reported that the 40 nm BPEI-AuNP inhibited gene expression of CYP1A2, CYP2C9, and CYP3A4 in human hepatocytes; ESD in HUVEC; and CYP1A1 in HPTC [7,8,9]. Epidemiology study demonstrated that CYP enzymes in liver tissue of HCC patient were substantially inhibited by the tumorigenic process at the molecular and the functional level [41].
Drug Uptake and Efflux Transporter Gene Expression Profiling
The development of multidrug resistance (MDR) by tumor cells is one of the main causes of cancer treatment failures [42, 43]. Integral membrane transporters-mediated decrease in drug uptake and increase in drug efflux including P-glycoprotein (P-gp) and breast cancer resistance protein (BCRP) is one of the major mechanisms of MDR.
Differential gene expression of drug efflux and uptake transporters in C3A cells exposed to 40 nm PEG-AuNP showed that a total of 14 genes of ABC transporters (↓12 and ↑2 genes) and a total of 21 genes of SLC transporters (↓21 and ↑0 genes) were substantially modulated at LC50 value (Figs. 6d and 7, Additional file 1:Table S7). In drug efflux transporters of ABC family, genes of multidrug resistance-associated protein (MRP3/ABCC3), MRP4 (ABCC4), and cholesterol efflux regulated protein (CERP/ABCA1) in basolateral membrane were downregulated (7.2- to 10.4-fold). The genes encoding P-gp (ABCB1), MRP2 (ABCC2), BCRP (ABCG2), and sterolin 2 (ABCG8) in canalicular efflux transporters were also suppressed (8.6- to 13.8-fold). In contrast, multidrug resistance (MDR4/ABCB4) in canalicular membrane and mitochondria ABC transporter (MTABC3/ABCB6) in the outer mitochondrial membrane were highly upregulated (9.8-fold and 5.8-fold, respectively). In drug uptake transporters, genes of copper transporter protein (CTR1/SLC31A1) and to a lesser degree organic anion transporting (OAT7/SLC22A9) were also inhibited (18- fold and 15-fold, respectively). These results support a recent study that the 40 nm BPEI-AuNP downregulate MDR3 in human hepatocytes but upregulates MRP3 in HUVEC indicating surface charge- and cell type-dependent interaction between AuNP and efflux transporters [7, 8]. Epidemiology study exhibited that a high expression of BCRP and a low expression of OCT3 occurred in HCC tumor, which was closely associated with the tumor progression and its size [44]. A previous study exhibited that P-gp inhibitor, verapamil enhanced cytotoxicity of glutathione-AuNP conjugated with doxorubicin in feline fibrosarcoma cell lines by increasing intracellular drug concentration [45]. The current study emphasizes that the mechanisms-derived information on the 40 nm PEG-AuNP identified a separate but still complementary action on mitochondrial fatty acid β-oxidation, TCA cycle and respiratory chain, drug efflux and uptake transporters, as well as CYP3A4 activity in C3A cells (Fig. 7). To the end, this will highlight AuNP interaction with key biological processes and its underlying molecular mechanism in HCC, which may be further implicated in the development of more effective therapeutic target in HCC treatment.

A schematic representation of the basic mechanisms of action of 40 nm PEG-AuNP in HCC treatment. Green bars (an inhibition) and pink triangles (an induction) indicate the 40 nm PEG-AuNP-modified biological markers and pathways. Gene ontology analysis is listed in Additional file 1:Tables S2–S7
To validate gene expression analysis from RT 2 array, the nine genes were selected for real-time PCR. In Additional file 1:Table S1, all nine genes were modulated at LC50 of the 40 nm PEG-AuNP. These transcriptional changes were consistent with those in gene expression analysis with PCR arrays (Fig. 6, Additional file 1:Tables S2–S7).
Conclusions
We have presented that cationic BPEI-, anionic LA-, or neutral PEG-AuNP interaction with human plasma protein corona (PC) caused the changes in D H , PDI, and z-potential of AuNP and further influenced cellular responses in C3A cells. All bare (no PC) 40 and 80 nm AuNP were cytotoxic to C3A cells besides the 80 nm LA-AuNP but PC completely ameliorated their cytotoxicities besides the 80 nm BPEI-AuNP. The 40 nm bare BPEI-AuNP showed the highest cellular uptake followed by the 40 nm PEG-AuNP and then the 40 nm LA-AuNP, whereas PC suppressed AuNP uptake besides the 80 nm PEG-AuNP. The 40 nm BPEI-AuNP caused biphasic responses of oxidative stress (pro- and antioxidant) in C3A cells, whereas the 40 nm PEG-AuNP was antioxidant. CYP3A4 activity was extensively suppressed by all bare AuNP, irrespective of size and surface charges, whereas PC substantially ameliorated its inhibitory effect on enzyme activity besides the 40 and 80 nm PEG-AuNP. Differentially expressed genes at LC50 value of 40 nm PEG-AuNP were mainly involved in mitochondrial fatty acid β-oxidation and to a lesser degree hepatic efflux/uptake transporters. The 40 nm PEG-AuNP inhibited three main enzymes in β-oxidation (acyl-CoA dehydrogenase, enoyl-CoA hydratase, and ketoacyl-CoA thiolase), other enzymes in TCA cycle, and the mitochondrial respiratory chain for ATP production. The 40 nm PEG-AuNP increased the expression of pro-apoptotic genes and decreased anti-apoptotic genes at LC50 value. A high level of antioxidants and a low level of pro-oxidants genes were observed in C3A cells exposed to 40 nm PEG-AuNP. In addition, genes of drug efflux and uptake transporters located in both basolateral and canalicular membrane were substantially modulated.
Afkortingen
- ANOVA:
-
One-way analysis of variance
- AuNP:
-
Gold nanoparticles bare:no PC
- BPEI:
-
Branched polyethylenimine
- CYP:
-
Cytochrome P450
- D H :
-
Hydrodynamic diameters
- DLS:
-
Dynamische lichtverstrooiing
- EDTA:
-
Ethylenediaminetetraacetic acid
- EMEM:
-
Eagle’s minimum essential medium
- HCC:
-
Human hepatocellular carcinoma
- HPTC:
-
Human renal proximal tubular cells
- HSD:
-
Tukey’s honest significant difference test
- HUVEC:
-
Human umbilical vein cells
- ICP-MS:
-
Inductively coupled plasma mass spectrometry
- LA:
-
Anionic lipoic acid
- LC50 :
-
Median lethal concentration
- MDR:
-
Multidrug resistance
- NP:
-
Nanodeeltjes
- PBS:
-
Phosphate-buffered saline
- PC:
-
Human plasma protein corona
- PDI:
-
Polydispersity index
- PEG:
-
Neutral polyethylene glycol
- RNAi:
-
RNA interference
- RNS:
-
Reactive nitrogen species
- ROS:
-
Reactieve zuurstofsoorten
- SO:
-
Superoxide
- TEM:
-
Transmissie-elektronenmicroscopie
- TFF:
-
Tangential flow filtration
Nanomaterialen
- Hotelonderhoud en de impact ervan op uw bedrijfsresultaten
- Gouden nanobiosensor gebaseerd op de gelokaliseerde oppervlakteplasmonresonantie kan menselijke brucellose diagnosticeren, wat een snelle en betaalbare methode introduceert
- Synthese en CO-oxidatieactiviteit van 1D gemengd binair oxide CeO2-LaO x ondersteunde gouden katalysatoren
- Vastestofverwarmingssynthese van poly (3,4-ethyleendioxythiofeen)/goud/grafeencomposiet en de toepassing ervan voor Amperometrische bepaling van nitriet en jodaat
- Bevordering van SH-SY5Y-celgroei door gouden nanodeeltjes gemodificeerd met 6-mercaptopurine en een neuron-penetrerend peptide
- Magnetic Gold Nanoparticle-Labeled Heparanase Monoclonal Antibody en de daaropvolgende toepassing voor Tumor Magnetic Resonance Imaging
- C60 Fullereen effecten op difenyl-N-(trichlooracetyl)-amidofosfaat interactie met DNA in Silico en zijn cytotoxische activiteit tegen menselijke leukemische cellijn in vitro
- Toxiciteitsbeoordeling van PEG-PCCL-nanodeeltjes en voorlopig onderzoek naar het antitumoreffect van Paclitaxel-loading
- Invloed van koolstofnanobuisjes en zijn derivaten op tumorcellen in vitro en biochemische parameters, cellulaire bloedsamenstelling in vivo
- Beoordeling van door gouden nanodeeltjes geremde cytochroom P450 3A4-activiteit en moleculaire mechanismen die ten grondslag liggen aan de cellulaire toxiciteit ervan in humane hepatocellulaire carcin…
- Fluorescerende neoglycoproteïne gouden nanoclusters:synthese en toepassingen in lectinedetectie bij planten en celbeeldvorming