


Frequentie-gemoduleerde golfdiëlektroforese van blaasjes en cellen:periodieke U-bochten bij de crossover-frequentie
Abstract
We hebben de diëlektroforetische kracht geformuleerd die wordt uitgeoefend op micro/nanodeeltjes bij de toepassing van frequentiegemoduleerde (FM) elektrische velden. Door het frequentiebereik van een FM-golf aan te passen om de crossover-frequentie f . te dekken X in het reële deel van de Clausius-Mossotti-factor, voorspelt onze theorie de omkering van de diëlektroforetische kracht telkens wanneer de momentane frequentie periodiek f doorloopt X . In feite hebben we periodieke U-bochten van blaasjes, leukemiecellen en rode bloedcellen waargenomen die FM-golf-diëlektroforese (FM-DEP) ondergaan. Onze theorie suggereert ook dat de video-tracking van de U-bochten als gevolg van FM-DEP beschikbaar is voor de behendige en nauwkeurige meting van f X . De FM-DEP-methode vereist een korte duur, minder dan 30 s, terwijl de FM-golf wordt toegepast om verschillende U-bochten te observeren, en de behendigheid bij het meten van f X is van groot nut voor niet alleen zoute celsuspensies, maar ook nanodeeltjes omdat de door het elektrisch veld geïnduceerde oplosmiddelstroom zoveel mogelijk wordt onderdrukt. De nauwkeurigheid van f X is geverifieerd met behulp van twee soorten experimenten. Eerst hebben we de aantrekkingskracht gemeten die wordt uitgeoefend op een enkel blaasje dat wisselstroom diëlektroforese (AC-DEP) ervaart bij verschillende frequenties van sinusoïdale elektrische velden. De frequentie-afhankelijkheid van de diëlektroforetische kracht levert f . op X als een karakteristieke frequentie waarmee de kracht verdwijnt. Het AC-DEP-resultaat van f . vergelijken X met die verkregen met de FM-DEP-methode, beide resultaten van f X bleken met elkaar samen te vallen. Ten tweede onderzochten we de geleidbaarheidsafhankelijkheden van f X voor drie soorten cellen door de omringende elektrolyten te veranderen. Op basis van de experimentele resultaten evalueerden we gelijktijdig zowel de cytoplasmatische geleidbaarheid als de membraancapaciteiten met behulp van een uitgebreide theorie over het single-shell-model van biologische cellen. Hoewel de cytoplasmatische geleidingsvermogens, vergelijkbaar voor deze cellen, iets lager waren dan het bereik van eerdere rapporten, kwamen de verkregen membraancapaciteiten goed overeen met die eerder in de literatuur werden gerapporteerd.
Achtergrond
De polariseerbaarheid van een elektrisch fenotype is voornamelijk te wijten aan het celmembraan en de cytoplasmatische elektrische eigenschappen die afhankelijk zijn van de frequentie van het aangelegde elektrische veld. Dienovereenkomstig kunnen individuele cellen worden geïdentificeerd door de verschillen in de diëlektrische spectra met behulp van niet-invasieve elektrische technieken. De elektrische technieken zijn momenteel competent voor het scheiden van cellen met bruikbare fenotypes van onbekende monsters [1-15]. Vergeleken met andere scheidingsmethoden bieden deze het grote voordeel dat celmodificatie door antilichamen of hechting aan vreemd materiaal niet nodig is, waardoor de kans op celbeschadiging of activering door deze sondes wordt vermeden [1–16]. De karakterisering van de cellulaire diëlektrische eigenschappen is voornamelijk uitgevoerd met behulp van ofwel impedantiespectroscopie [10, 12, 13] of wisselstroom (AC) elektrokinetiek zoals diëlektroforese (DEP), lopende-golf DEP (twDEP) en elektrorotatie [1, 9, 15]. Onder hen richten we ons op het uitbreiden van de AC-DEP-methode om een nieuwe methode voor diëlektrische karakterisering te ontwikkelen met behulp van frequentiegemoduleerde (FM) golven in plaats van AC-velden.
Over het algemeen vindt de DEP plaats in een elektrisch veldgradiënt die een elektrokinetische kracht creëert die wordt uitgeoefend op elk polariseerbaar object, geladen of neutraal, in de richting die niet alleen wordt bepaald door de gradiëntvector, maar ook door het reële deel van de Clausius-Mossotti ( CM)-factor [1–15, 17–21]. We beschouwen bijvoorbeeld de DEP-kracht die wordt geïnduceerd door het AC-elektrische veld E AC (r ,t ) waarvan de ruimte-tijdafhankelijkheid wordt uitgedrukt als E AC (r ,t )=A (r ) cosθ AC (r ,t ) met behulp van de amplitudevector A (r ) en de fase θ AC (r ,t ). De AC-DEP-kracht wordt gegenereerd door de ruimtelijke gradiënt van de amplitude (d.w.z. ∇A ) vermenigvuldigd met het reële deel van de CM-factor, zoals hierboven vermeld, terwijl de ruimtelijke gradiënt van de fase (d.w.z. ∇θ AC ) vermenigvuldigd met het denkbeeldige deel van de CM-factor creëert de kracht van twDEP of elektrorotatie, wat daarom aanvullende informatie geeft aan de AC-DEP-methode in termen van de diëlektrische karakterisering [9, 15, 20, 21].
In deze brief willen we de DEP-kracht die wordt veroorzaakt door een FM-veld formuleren en de AC- en FM-DEP-methoden vergelijken, zodat noch het AC- noch het FM-veld rekening houdt met de ruimtelijke afhankelijkheid van de fase; daarom zullen we θ . instellen AC (t )=2π f AC t in verhouding tot de toegepaste frequentie f AC . Een belangrijk kenmerk van de AC-DEP is dat zowel de krachtrichting als de sterkte ervan afhangt van f AC . Het meest opvallende is dat de krachtrichting wordt omgekeerd bij crossover-frequentie f AC =f X vanwege de verandering in teken van het reële deel van de CM-factor, die beschikbaar is gebleken voor de diëlektrische karakterisering met behulp van AC-DEP [1–15].
De frequentie-afhankelijkheid van de AC-DEP-kracht heeft ook de volgende manipulaties mogelijk gemaakt [1-15, 22-31]:elektrisch regelbare vangst, focussering en translatie van colloïdale deeltjes, evenals de fractionering en karakterisering van levende en/of dode cellen. Conventionele systemen voor de diëlektroforetische assemblage en/of manipulatie van colloïdale deeltjes hebben vaak gebruik gemaakt van microgefabriceerde elektroden waartussen het AC-elektrische veld is toegepast op colloïdale suspensies, profiterend van recente snelle vorderingen in de fabricage van geïntegreerde halfgeleiderinrichtingen [24-30] . Deze technologie, die contactloze manipulatie biedt, wordt momenteel geïntegreerd met een verscheidenheid aan lab-on-a-chip-systemen die het voordeel bieden van nauwkeurige en herhaalbare hantering. Desalniettemin zijn de on-chip elektroden die plekken met hoge intensiteit in AC-velden creëren niet in staat om hun posities onafhankelijk van de monsterhouder te veranderen, in tegenstelling tot de laserfocus die vrij kan worden gepositioneerd in de optische manipulatie. Uit de beperking van on-chip-systemen volgt dat de vorige DEP-methoden enige moeilijkheid en complexiteit hebben opgeleverd bij het uitvoeren van de soorten bewerkingen waarvoor optische pincetten geschikt zijn. Een mogelijke methode om deze moeilijkheden te overwinnen is optische beeldgestuurde DEP [32].
Hier hebben we, als een eenvoudiger alternatief, een van de elektronische pincettentechnieken [22, 23, 33-38] aangenomen voor on-demand diëlektroforetische assemblage en / of manipulatie zonder een optisch apparaat (zie figuur 1). Zoals te zien is in figuur 1, gebruikt ons plug-in-systeem een paar micro-elektrodenaalden die worden bestuurd door micromanipulatoren voor het aanbrengen van de externe elektrische velden in een colloïdale suspensie. De elektrodesondes waren niet vast maar beweegbaar in colloïdale suspensies vanwege hun plug-in-stijl. Er blijft echter een belangrijke vereiste voor het praktische gebruik van diëlektrische karakterisering:de duur dat een elektrisch veld wordt aangelegd op de cellen die worden omgeven door zoute elektrolyten, moet worden geminimaliseerd. De AC-DEP-methode omvat bijvoorbeeld het gebruik van in elkaar grijpende kamachtige elektroden die zijn ingebed in een microfluïdisch systeem, zodat de AC-velden van verschillende frequenties gelijktijdig kunnen worden toegepast in een celsuspensie [24-30]. Hoewel dergelijke verfijnde on-chip-systemen relevant zijn bevonden voor de diëlektrische karakterisering, is de multi-elektrodepaartechniek niet toepasbaar op het systeem met één elektrodepaar dat vaak is gebruikt in de elektronische pincettechnieken [22, 23, 33-38] .

Experimentele opstelling. Een schema van het diëlektroforetische manipulatiesysteem dat het AC- of FM-elektrische veld illustreert dat op een doeldeeltje wordt toegepast via een paar elektrodenaalden die worden bestuurd door micromanipulatoren met patch-clamp
Om gelijktijdige multifrequentiemetingen uit te voeren met behulp van het systeem met één elektrodepaar (Fig. 1), moet de verandering in het aangelegde elektrische veld worden onderzocht. In deze brief gaan we in op de beschikbaarheid van in de tijd variërende DEP als gevolg van een FM-golf (FM-DEP) van de volgende vorm:
$$ \boldsymbol{E}(\boldsymbol{r},t) =\boldsymbol{A}(\boldsymbol{r})\cos\theta(t), $$ (1)waarbij de fase θ (t ) van de FM-golf is gerelateerd aan de momentane frequentie f (t ) als 2π f (t )=d θ (t )/d t en
$$ f(t)=f_{c}+\Delta f\cos\left(2\pi f_{m}t \right), $$ (2)met f m aanduiding van de modulatiefrequentie. We gebruiken de breedband FM die voldoet aan dat Δ f /f m ≫1, zodat de voorwaarden van f m /f (t ), f m /f c , f m /Δ f ≪1 zal worden aangeduid als de breedbandlimiet (WBL) in de theoretische formulering die hieronder wordt gegeven.
In deze brief wordt bijzondere aandacht besteed aan de relatie tussen de karakteristieke frequentie van f X en het traject van FM-DEP. In de volgende sectie beschrijven we zowel de gebruikte materialen als details van het plug-in systeem voor het induceren van FM-DEP. Het derde deel geeft de resultaten en de discussie die uit vier delen bestaat. Eerst onderzoeken we de details van herhalende U-bochten van een enkele leukemiecel door het heen en weer gaande traject te kwantificeren, waarvan de periodiciteit wordt verklaard door de modulatiefrequentie f m , of de periodieke oscillatie van f (t ) gegeven door Vgl. (2). Vervolgens leggen we het heen en weer gaande traject theoretisch uit door de in de tijd variërende diëlektroforetische kracht af te leiden die moduleert volgens de momentane frequentie f (t ) van het FM-veld dat voldoet aan de WBL-voorwaarde. De verkregen vorm van de diëlektroforetische kracht levert de vergelijking die de f . bepaalt X van de waargenomen U-bochten. Ten derde meten we de grootte van de diëlektroforetische kracht op een multilamellaire vesikel (MLV) die aan een elektrodenaald was bevestigd vanwege de aantrekking van de AC-DEP. De frequentie-afhankelijkheid van kracht werd aangepast met behulp van de spectrale vergelijking die werd bepaald uit het reële deel van de CM-factor, zodat f X werd bepaald als de karakteristieke frequentie waarmee de aantrekkingskracht als gevolg van AC-DEP verdwijnt. Omdat de FM-DEP-methode ook f . geeft X door het heen en weer gaande traject van een MLV te analyseren, beoordelen we de mate van coïncidentie tussen de crossover-frequenties die zijn geëvalueerd op basis van AC- en FM-DEP's. Ten slotte werden zowel de cytoplasmatische geleidbaarheid als de membraancapaciteiten van drie soorten cellen geëvalueerd uit f X als een toenemende functie van de geleidbaarheid van de oplossing, en de verkregen waarden werden vergeleken met die gerapporteerd in de literatuur.
Methoden
Materialen
Voor het bereiden van multilamellaire blaasjes (MLV's), gebruikten we 1,2-dioleoyl-sn-glycero-3-fosfatidylcholine (DOPC) als lipiden, gekocht bij Avanti Polar Lipids. De MLV's werden verkregen door de volgende procedure. De DOPC (1 ml, 20 mM) opgelost in chloroform/methanol (2:1 v /v ) werd gedroogd met N2 gas, en het oplosmiddel werd gedurende meer dan 12 uur onder vacuüm volledig verwijderd. De dunne film die door de verdamping op het glazen flesje was afgezet, werd gerehydrateerd met gedeïoniseerd water en enkele uren bij 25 °C geïncubeerd.
Twee cellijnen die in de experimenten werden gebruikt, waren JKT-beta-del van de lijn van menselijke T-celleukemie (TL) en CCRF-SB van de lijn van menselijke B-celleukemie (BL). Beide soorten TL- en BL-cellen werden gebruikt na 1 week incubatie in een bevochtigde incubator die 5 % bevat CO2 op 37, zodat we de celconcentraties hadden binnen het bereik van 0,5×10 6 tot 1×10 6 cellen/ml. Het RPMI 1640-medium voor de celkweek werd aangevuld met 10% foetaal bovien serum en 100 mM natriumpyruvaat. De cellen werden gesedimenteerd door centrifugatie bij 370g gedurende 3 min tweemaal, zodat de cellen zuiver kunnen worden geresuspendeerd in 1 ml van het RPMI 1640 medium voorafgaand aan het pipetteren. De verkregen celsuspensies werden verder verdund met behulp van de isotone 200 mM sucrose-oplossing om het oplosmiddel voor te bereiden met een vereiste geleidbaarheid.
We gebruikten ook menselijke rode bloed (RB) cellen gedispergeerd in de volgende suspensies. Vers afgenomen volbloedmonsters werden verkregen van gezonde vrijwilligers van begin twintig. De cellen, gesuspendeerd in een mengsel van het RPMI 1640-medium en hematocriet van 3,1%, werden verdund met behulp van de isotone 200 mM sucrose-oplossing om het oplosmiddel voor te bereiden met een vereiste geleidbaarheid, evenals de bovenstaande leukemiecellen. Alle diëlektroforetische experimenten met menselijke RB-cellen waren binnen 10 minuten na het afnemen van de volbloedmonsters voltooid.
Experimentele setup
De geleidbaarheid van de celsuspensies werd gemeten met behulp van een geleidbaarheidsmeter (SevenMulti, Mettler-Toledo, Columbus, OH, VS). Een schema van het gebruikte plug-insysteem wordt getoond in Fig. 1. Een extern elektrisch veld met een AC- of FM-golf werd aangelegd via een willekeurige golfvormgenerator (Agilent 33220A, Agilent Technologies, Santa Clara, CA, VS) met een stroom versterker (F30PV, FLC Electronics, Partille, Zweden) waarop plug-in-type micro-elektroden waren aangesloten. De micro-elektroden bestonden uit wolfraamnaalden met een puntdiameter van 0,5 μ m die onafhankelijk werden bestuurd door twee sets patch-clamp-micromanipulatoren (NMN-21, Narishige, Setagaya-ku, Tokyo, Japan). In alle volgende experimenten handhaafden we de puntscheiding op 100 μ m bij het toepassen van de externe velden op de bovenstaande suspensies, en de maximale grootte werd ingesteld op 0,5 kV/cm. Het naaldpaar werd ingebracht in een monsterdruppel gemonteerd op de omgekeerde optische microscoop (TE2000-U, Nikon, Minato-ku, Tokyo, Japan), en de optische microfoto's werden verkregen met behulp van een CCD-camera (Retiga Exi, QImaging, Surrey, Brits Columbia, Canada) met een framesnelheid van 25 fps; overigens werd bevestigd dat de frequentieresolutie van FM-golven als gevolg van de framesnelheid altijd binnen de foutbalken voor elke gegevens lag. Een 50- μ Een druppel suspensie werd op het monsterplatform van de omgekeerde optische microscoop gemonteerd, waarvan de temperatuur met een warmteregelaar op 25 °C werd gehouden.
De plug-in techniek stelt het eenvoudige systeem in staat om verschillende contactloze manipulaties van een enkele cel uit te voeren, zoals het zonder enig contact in een smal kanaal duwen en het in de gewenste richting oriënteren. Hoewel het vaak nodig is om de cellen in een isotone oplossing met zout te behandelen, is het het gemakkelijkst om de bovengenoemde DEP-manipulaties van cellen omgeven door gedeïoniseerd water uit te voeren. In Aanvullend bestand 1:Films S1 tot S3 induceerde het plug-insysteem de AC-DEP van diatomeeëncellen gesuspendeerd in gedeïoniseerd water. We kunnen uit aanvullend bestand 1:Films S1 tot S3 zien dat een anisotrope diatomeeëncel gedispergeerd in zoutvrij water als een post-it-tag werd gemanipuleerd door een paar micro-elektroden waartussen het elektrische wisselstroomveld (1 kV/cm) werd aangebracht . De contactloze operaties bestaan uit drie stappen:(i) een doelcel werd eerst parallel gedraaid aan een glazen wand die positief geladen was door de combinatie van dipooluitlijning met een frequentie van 30 kHz en positieverandering van elke micro-elektrode (aanvullend bestand 1:film S1 ), (ii) we hebben vervolgens de frequentie gewijzigd in 100 kHz om deze naar de muur te duwen om de verzoekcel elektrostatisch met negatieve ladingen op het glasoppervlak te fixeren (Aanvullend bestand 1:Film S2), en (iii) de AC-frequentie werd aangepast tot 20 MHz om de AC-DEP in de tegenovergestelde richting te induceren, zodat de elektrostatisch bevestigde cel eruit kon worden getrokken (Aanvullend bestand 1:Film S3).
Resultaten en discussie
Experimentele observatie van een leukemiecel die FM-DEP ervaart
Met onze plug-in micro-elektroden (zie Fig. 1) kan het elektrische veld worden toegepast op de deeltjes die ver boven het monstersubstraat zweven, wat van praktisch nut is voor het selecteren van geschikte cellen. Aanvullend bestand 1:Film S4 laat bijvoorbeeld zien dat het micro-elektrodepaar werd bestuurd om een zwevende driehoekige diatomeeëncel te naderen waarop we het AC-elektrische veld hebben toegepast met zijn frequentiesprong tussen 100 en 500 kHz met intervallen van 0,5 s. In Aanvullend bestand 1:Film S4 zien we de driehoekige cel stuiteren op een micro-elektrode vanwege het springen van de frequentie als een voorlopig resultaat voorafgaand aan de volgende manipulatie met behulp van de FM-DEP.
Aanvullend bestand 1:Films S5 en S6 tonen typisch gedrag van verschillende TL-cellen die de FM-DEP ervaren, die vergelijkbaar zijn met die van zoogdiercellen die worden gemanipuleerd door een elektronisch pincet met behulp van een enkele elektrode AC-DEP [36]. Figuur 2 toont een van de periodieke trajecten met behulp van de 3D-plot van (x ,j ) langs de t as, waarbij een relatieve coördinaat van (x ,j ) wordt toegewezen aan de tijdelijke celpositie met de oorsprong van (0, 0) op een specifiek punt op een micro-elektrodenaald voor het extraheren van de cel-elektrodeconfiguratie. Terwijl de x as vertegenwoordigt de raaklijn aan het elektrodeoppervlak bij (0, 0), de y as, loodrecht op de raaklijn, weerspiegelt voornamelijk de projectie van de hieronder toegelichte periodieke U-bochten. In Fig. 2 hebben we een zwevende TL-cel geselecteerd waarop we het FM-elektrische veld met zijn modulatiefrequentie f hebben toegepast m ingesteld op f m =0,25 Hz in het bereik van 200 kHz ≤f (t )≤ 3 MHz. Omdat we die Δ . hebben f /f m , f (t )/f m <10 −5 , de WBL-conditie geldt eigenlijk, zoals vermeld na Vgl. (2).

3D-traject van een gerichte TL-cel. Periodieke U-bochten als gevolg van de frequentiemodulatie worden aangetoond voor de TL-cel die de FM-DEP ondergaat
Uit aanvullend bestand 1:Films S5 en S6 en uit figuur 2 blijkt dat het periodieke traject wordt gevormd door drie delen van het verlaten, naderen en blijven op de micro-elektrode:(i) de cel verlaat de micro-elektrode, (ii) het nadert de micro-elektrode na het maken van een U-bocht, en (iii) het blijft op het oppervlak van de micro-elektrode. De cel kan vaak niet terugkeren naar dezelfde positie op het oppervlak van de micro-elektrode vanwege de oplosmiddelstroom, die niet alleen wordt waargenomen in aanvullend bestand 1:film S6, maar ook wordt weergegeven door de U-bochten waarbij de cel migreert in de x richting in Fig. 2. Ondanks de interferentie met de oplosmiddelstroom, is het mogelijk om de momenten te onderscheiden waarop de cel respectievelijk het micro-elektrodeoppervlak begint te verlaten en de U-bocht in het periodieke traject maakt. Dienovereenkomstig kunnen we uit Fig. 2 zien dat deze U-bochten worden herhaald met intervallen van 4 s in overeenstemming met de modulatiefrequentie van 0,25 Hz, of de periode van 4 s van de momentane frequentie f (t ).
Theoretisch onderzoek naar de FM-DEP
Om de experimentele banen te verklaren, inclusief de periodieke U-bochten, beschouwen we een bolvormig object als een vereenvoudigd model van een enkele cel, waaraan een willekeurig in de tijd variërend elektrisch veld E (r ,t ) is toegepast. Figuur 3 toont een schema van de tijdsafhankelijke DEP-kracht die op een bolvormig object inwerkt [9]. Zoals getoond in Fig. 3, worden de permittiviteit en geleidbaarheid in een bolvormig object weergegeven door ε in en σ in , respectievelijk, en het subscript "out", zoals ε uit en σ uit , geeft de buitenkant aan. In het algemeen, F DEP (r ,t ) is gerelateerd aan het geïnduceerde dipoolmoment p (r ,t ) als [17-19]
$$\begin{array}{@{}rcl@{}} \boldsymbol{F}_{\text{DEP}}(\boldsymbol{r},t)&=&\left\{\boldsymbol{p} (\boldsymbol{r},t)\cdot \nabla\right\}\boldsymbol{E}(\boldsymbol{r},t), \end{array} $$ (3)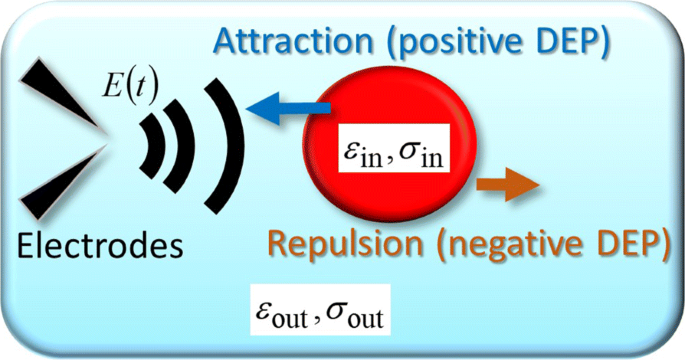
Theoretisch model. Schematische weergave van de FM-DEP-kracht die wordt uitgeoefend op een cel die is gemodelleerd als een homogeen bolmodel met de permittiviteit en geleidbaarheid van ε in en σ in , respectievelijk. De bol is omgeven door een elektrolytmedium met zijn permittiviteit en geleidbaarheid van ε uit en σ uit , respectievelijk. Het homogene bolmodel is de vereenvoudiging van het bolvormige enkele schaalmodel dat de cel beschouwt als een uitgesmeerd cytoplasma omgeven door een membraan [9]
$$\begin{array}{@{}rcl@{}} \boldsymbol{p}(\boldsymbol{r},t)&=&4\pi R^{3}\epsilon_{\text{out}}K_ {H}\left\{\boldsymbol{E}(\boldsymbol{r},t)+\frac{\tau}{\Delta\tau} \widetilde{\boldsymbol{E}}(\boldsymbol{r}, t)\right\}, \end{array} $$ (4) $$\begin{array}{@{}rcl@{}} \widetilde{\boldsymbol{E}}(\boldsymbol{r},t )&=&\frac{1}{\tau}\int_{0}^{tds}\,\boldsymbol{E}(\boldsymbol{r},ts)e^{-s/\tau}, \end {array} $$ (5)waar K H en Δ τ worden als volgt gedefinieerd:K H =(ε in −ε uit )/(ε in +2ε uit ), en \(\Delta \tau ^{-1}=\tau _{0}^{-1}-\tau ^{-1}\) met de straal R van het bolvormige object en twee karakteristieke tijden van τ 0 =(ε in −ε uit )/(σ in −σ uit ) en τ =(ε in +2ε uit )/(σ in +2σ uit ).
Vervanging van het elektrische wisselstroomveld E AC (r ,t )=A (r ) cos(2π f AC t ) in vergelijkingen. (3) tot (5), we verkrijgen de gemiddelde DEP-kracht <F DEP (r ,t )> dat is gemiddeld over cycli van het AC-veld [9, 15, 20]:
$$\begin{array}{@{}rcl@{}} \left<\boldsymbol{F}_{\text{DEP}}\right>&=&4\pi R^{3}\epsilon_{\text {out}}K_{H}\left[ \left<\boldsymbol{E}\cdot \nabla\boldsymbol{E}\right>+\frac{\tau}{\Delta\tau}\left<\widetilde{ \boldsymbol{E}}\cdot \nabla\boldsymbol{E}\right>\right]\\ &=&\chi(f_{\text{AC}})\nabla\boldsymbol{A}^{2}_ {\text{RMS}}, \end{array} $$ (6)waar A RMS geeft de RMS-vector (root mean squared) aan die voldoet aan \(\boldsymbol {A}_{\text {RMS}}^{2}=\boldsymbol {A}^{2}/2\), en χ (f AC )≡2π R 3 ε uit Re[K (f AC )] hangt af van de toegepaste frequentie f AC vanwege Re[K (f AC )], het reële deel van de CM-factor [9, 15, 20]:
$$\begin{array}{@{}rcl@{}} \chi(f_{\text{AC}})=\frac{2\pi R^{3}\epsilon_{\text{out}}} {1+(2\pi f_{\text{AC}}\tau)^{2}} \left\{ K_{L}+(2\pi f_{\text{AC}}\tau)^{2 }K_{H} \right\}, \end{array} $$ (7)waar K L =(σ in −σ uit )/(σ in +2σ uit ) en K H , hierboven gedefinieerd, komen overeen met de echte CM-waarden in respectievelijk de lage en hoge frequentiegrenzen, en deze grenswaarden, K L en K H , moeten tegengestelde tekens hebben zodat f X gedefinieerd door χ (f X )=0 kan bestaan [9, 15, 20].
Vergelijkingen (6) en (7) geven aan dat het AC elektrische veld de DEP-kracht creëert waarvan de richting afhangt van de toegepaste frequentie f AC via χ (f AC ) gegeven door Vgl. (7), waarin de stuiterende diatomeeëncel in Aanvullend bestand 1:Film S4 als volgt wordt uitgelegd (zie ook Fig. 3). Wanneer de toegepaste frequentie het plusteken geeft van het reële deel van de CM-factor (d.w.z. χ (f AC )>0), kunnen we cellen waarnemen die worden aangetrokken door de uiteinden van de elektrodenaald (de positieve DEP) waarop de sterkte van het AC-veld, aangebracht via een paar elektrodenaalden, het grootst is. Het teken van de reële CM-factor kan worden omgekeerd naar het negatieve bij f X , de verdwijnfrequentie van de echte CM-factor (d.w.z. χ (f X )=0), waarbij we de diëlektroforetische kracht nul hebben zoals gevonden in Vgl. (6). In het negatieve teken van de CM-factor (d.w.z. χ (f AC )<0), worden individuele colloïden afgestoten van het elektrodenaaldpaar (de negatieve DEP). De driehoekige diatomeeëncel in aanvullend bestand 1:film S2 stuiterde vanwege de tegengestelde richting van de AC-DEP's die werden veroorzaakt door de AC-velden met hun frequenties van 100 en 500 kHz; combineren vgl. (6) en de waargenomen diëlektroforetische richtingen, vinden we dat χ (100 kHz)>0 en χ (500 kHz)<0.
Vervolgens beschouwen we de FM-DEP door de fase die wordt gegeven door Vgl. (1) en (2) in Vgl. (3) tot (5). Zoals bewezen in aanvullend bestand 2, valideert de WBL-conditie van de FM-golf de geschatte vorm van de integratie in Vgl. (5), waardoor
$$ \left<\widetilde{\boldsymbol{E}}\cdot \nabla\boldsymbol{E}\right>=\frac{1}{1+\{2\pi f(t)\tau\}^{ 2}}\left(\frac{\nabla\boldsymbol{A}^{2}_{\text{RMS}}}{2} \right), $$ (8)die dezelfde vorm krijgt als die van de AC-DEP wanneer de tijdsafhankelijke frequentie f (t ) wordt vervangen door een constante frequentie van f AC . We verkrijgen dus de beperkende vorm van de gemiddelde DEP-kracht <F DEP (r ,t )> dat is gemiddeld over cycli van θ (t ) in het FM-veld (zie Vgl. (A1), (A13) en (A14) in aanvullend bestand 2):
$$\begin{array}{@{}rcl@{}} \left<\boldsymbol{F}_{\text{DEP}}(\boldsymbol{r},t)\right>=\chi\{f (t)\}\nabla\boldsymbol{A}^{2}_{\text{RMS}}, \end{array} $$ (9)van een soortgelijke vorm als Vgl. (6) voor de AC-DEP. Het verschil is of de coëfficiënt van χ {f (t )} hangt af van t via f (t ), die cyclisch verandert volgens de frequentiemodulatie met de periode van T m =1/f m .
Gebaseerd op de eenvoudige uitdrukking (9) van de FM-DEP, illustreren we met Fig. 4 het mechanisme van de bovenstaande U-bochten als gevolg van de FM-golf. Figuur 4 toont een schema van de DEP die wordt geïnduceerd door de FM-golf in de WBL wanneer het bereik van f (t ) dekt de crossover-frequentie f X zodat f c −Δ f ≤f X ≤f c +Δ f . In figuur 4 wordt verondersteld dat de frequentie-afhankelijkheid van het reële deel van de CM-factor, of χ {f (t )}, geeft de alternatieve tekenwijzigingen als volgt:minteken (χ {f (t )}<0) voor f (t )<f X en plusteken (χ {f (t )}>0) voor f (t )>f X , wat het geval is met onze experimenten. De vorige periode die voldoet aan f (t )<f X heeft een tijdsduur, terwijl de laatste f (t )>f X is behouden tijdens de rust van de periode:één cyclus wordt ingedeeld in twee perioden die respectievelijk rood en blauw zijn gemarkeerd in Fig. 4.

Krachtrichting gerelateerd aan de frequentiemodulatie. Een illustratie van de periodieke U-bochten als gevolg van de FM-golf met zijn tijdsafhankelijke frequentie van f (t ) met een crossover-frequentie f X
Net als bij de AC-DEP, Vgl. (9) houdt in dat het minteken (χ {f (t )}<0) creëert een afstotende DEP-kracht tussen de cel en de micro-elektroden terwijl wordt voldaan aan dat f (t )<f X . Hierdoor verlaat de cel het gebied rond de naaldpunten van de micro-elektrode waartussen de grootte van het elektrische veld het grootst is:de cel ervaart de negatieve DEP tijdens de roodperiode van Δ t n in Fig. 4. Op het moment t X als een oplossing van f (t X )=f X , χ (f ) verdwijnt, gevolgd door het teken verandert in χ (f )>0 while f (t )>f X , en dienovereenkomstig wordt de DEP-kracht omgeschakeld naar de aantrekkende kracht bij t X . Na het maken van een U-bocht bij t X als gevolg van de omkering in de richting van de DEP-kracht, begint de beoogde cel de micro-elektrode te naderen die in de tegenovergestelde richting migreert en wordt uiteindelijk gevangen tussen de uiteinden van de elektrodenaalden of vastgemaakt aan een van de elektroden:de cel ervaart de positieve DEP tijdens de blauwe periode van Δ t p in Fig. 4. Figuur 4 geeft aan dat de cyclus van het verlaten, naderen en blijven op de micro-elektrode herhaald moet worden met de modulatieperiode van T m , in overeenstemming met Fig. 2:Δ t n +Δ t p =T m . Het diëlektroforetische mechanisme afgebeeld in Fig. 4 kan dus de periodieke U-bochten verklaren die zijn waargenomen in Aanvullend bestand 1:Films S5 en S6 evenals Fig. 2.
Laten we eens kijken naar de periodieke oplossing van de vergelijking, f (t X )=f X . Zoals te zien is in Fig. 4, t X wordt uitgedrukt als t X =n T m +0.5Δ t p =n T m +0.5(T m −Δ t n ) met een geheel getal van n =0, ±1, ±2,⋯, wat verder luidt als
$$ 2\pi f_{m} t_{X}=(2n+1)\pi-\pi f_{m}\Delta t_{n}. $$ (10)Vervanging van Eq. (10) in Vgl. (2), we hebben voor n =0 dat
$$ f_{X}=f_{c}-\Delta f\cos\left(\pi f_{m}\Delta t_{n}\right), $$ (11)verduidelijken dat de FM-DEP-methode de crossover-frequentie bepaalt als de tijdsduur Δ t n van het verlaten van de micro-elektrode tot het maken van de U-bocht kan nauwkeurig worden gemeten.
Vergelijking van crossover-frequenties van een enkele MLV bepaald op basis van de FM- en AC-DEP's
We onderzochten de experimentele nauwkeurigheid van Vgl. (11). Experimenteel is het vaak nodig dat de biologische cellen in een elektrolyt worden gedispergeerd. Voor MLV's is het gebruik van gedeïoniseerd water echter toegestaan tijdens het bereidingsproces van rehydratatie en verdunning. We hebben dus de zoutvrije MLV-suspensie gebruikt om de crossover-frequenties te vergelijken die zijn bepaald op basis van zowel AC- als FM-DEP's.
De diëlektroforetische U-bochten van een gerichte MLV werden veroorzaakt door de FM-golf in het bereik van 10 kHz ≤f (t )≤ 50 kHz (d.w.z. f c =30 kHz en Δ f =20 kHz) met een instelling die f m =0,1 Hz, en dienovereenkomstig heeft de FM-DEP een periode van 10 s. In de experimenten duurt het minder dan 30 s om een paar U-bochten van de beoogde MLV te observeren van het verlaten tot het naderen van micro-elektroden. Uit het traject hebben we de gemiddelde vertrektijd verkregen die \(\overline {\Delta t_{n}}=5.8\pm 0.2\) s. Omdat de WBL-voorwaarde van toepassing is op het huidige experiment dat voldoet aan de voorwaarde dat f m /Δ f /f m , f , m /f (t )<10 −5 , werd de crossover-frequentie beoordeeld als f X =35±1 kHz door \(\overline {\Delta t_{n}}=5.8\pm 0.2\) s te vervangen door Eq. (11).
For comparison, we made use of the programmable manipulator in the AC-DEP method that tries to evaluate the crossover frequency of the same targeted MLV to which the sinusoidal electric field with a frequency in the range of 30 to 100 kHz was applied via the electrode needle pair for inducing the AC-DEP. Because the programmable manipulator carries the electrode needle pair at a constant speed in one direction, we can measure the dielectrophoretic force similarly to the laser-trapping experiments [39]. Attaching the MLV on an electrode tip that undergoes uniform linear motion, not only the AC-DEP force but also the hydrodynamic force caused by the one-dimensional motion are exerted on the MLV. With the gradual increase of electrode velocity, F DEP eventually becomes smaller than the hydrodynamic force. As a result, the MLV initially attached to the moving electrode, owing to the DEP attraction, is desorbed by the hydrodynamic force. Defining the critical value, v c , by the maximum velocity value of the microelectrode pair prior to the desorption, the force balance equation between the DEP and hydrodynamic forces reads [39]
$$ F_{\text{DEP}}(f_{\text{AC}})=6\pi\eta R v_{c}, $$ (12)waar F DEP (f AC )e ≡<F DEP> with the unit vector e defined by \(\boldsymbol {e}=\nabla {\boldsymbol {A}}^{2}_{\text {RMS}}/|\nabla {\boldsymbol {A}}^{2}_{\text {RMS}}|\), η the water viscosity at 25 °C and 2R the diameter of the MLV.
Additional file 1:Movies S7 and S8 demonstrates the force measurement using the above AC-DEP method at the applied frequency of f AC =60 kHz. In Additional file 1:Movie S7, the velocity of the electrode pair controlled by the programmed manipulator is 110 μ m/s, which is lower than v c ; therefore, the MLV remains attached to one part of the electrode pair owing to the dielectrophoretic attraction. Additional file 1:Movie S8, on the other hand, shows the higher electrode speed of 120 μ m/s, under which the dielectrophoretic force becomes smaller than the hydrodynamic force that is exerted on the MLV, thereby desorbing the MLV from the electrode. Accordingly, v c is evaluated to be 110 μ m/s ≤v c ≤ 120 μ m/s, and we can calculate F DEP (60 kHz) using Eq. (12).
We can determine f X from the experimental results of F DEP at various external frequencies. Figure 5 shows the frequency dependence of F DEP , indicating that the DEP force experienced by the MLVs was reduced by lowering the applied frequency. It is found from Eqs. (6) and (7) that the fitting function of F DEP (f AC ) can be expressed as
$$ F_{\text{DEP}}(f_{\text{AC}})=\frac{L+(2\pi f_{\text{AC}}\tau)^{2}H}{1+(2\pi f_{\text{AC}}\tau)^{2}}, $$ (13)
Frequency dependence of F DEP . The FM-DEP force (F DEP ) as a function of external frequency (f AC ) of applied AC field where F DEP has been evaluated from Eq. (12), the balance equation between the FM-DEP and hydrodynamic forces exerted on a single MLV. It can be seen that F DEP is increased and saturated as f AC is higher, reflecting a typical behavior of the relaxation spectrum of the real CM factor. The solid line represents the best-fit result of Eq. (13)
implying that
$$ f_{X}=\frac{1}{2\pi\tau}\sqrt{-\frac{L}{H}}. $$ (14)Equation (13) is depicted by the solid line in Fig. 5 that has been fitted to the experimental data using the best-fit results of three parameters:L =−21.02 pN, H =19.03 pN, and τ =4.9 μ s. Substituting these results into Eq. (14), we evaluate that f X =34.15 kHz, which coincides with the result of f X =35±1 kHz evaluated from the FM-DEP method. The FM-DEP method is thus validated in terms of the consistency with the direct force measurement using the AC-DEP method.
Conductivity Dependencies of the Crossover Frequencies for Biological Cells
Let us return to the dielectrophoretic U-turns of biological cells mentioned in Fig. 2 to assess the practical reliability of the crossover frequencies when the FM-DEP method is applied to cell suspensions. Recently, an elaborate theory [40] has investigated, in more detail than before, the relationship between the homogeneous sphere model (see Fig. 3) and the single-shell model where the inner structure of cell is represented by a smeared-out cytoplasm surrounded by a membrane. As a result, the relation between f X and the suspension conductivity σ uit has been formulated using radius R of a cell, membrane capacitance C m , and cytoplasmic conductivity σ cyt [40]:
$$ f_{X}=\frac{1}{\sqrt{2}\pi {RC}_{m}}\left(\sigma_{\text{out}}-\frac{1}{2\sigma_{\text{cyt}}} \sigma_{\text{out}}^{2} \right)+f_{X0}, $$ (15)waar f X 0 is the extrapolated value to the crossover frequency at σ =0 mS/m and will be treated as a fitting parameter herein. The elaborate treatment adds the squared term, the second term on the right hand side of Eq. (15), to the conventional linear relation which has mainly been used for evaluating C m from f X [40–45]. Theoretically, it has still been claimed [40] that Eq. (15) is valid within a lower range of σ uit such that σ uit <10 mS/m; however, it should be better to include the squared term in the evaluation of C m , considering that our range of σ uit is relatively high compared with previous results in the range of 10 mS/m ≤σ uit ≤ 100 mS/m [40–45]. Hence, we determined σ cyt as well as C m from fitting Eq. (15) to the experimental results of f X as an increasing function of σ uit .
There are three kinds of biological cell used:TL and BL cells of human leukemia and RB cells of three human volunteers. In all the experiments using any species of cell, the conductivities were within the range of 60 to 160 mS/m, and the modulation frequency was set to be 0.25 Hz. Regarding the instantaneous frequency, most of the experiments adopted the range from 100 to 1.5 MHz (i.e., f c =800 kHz and Δ f =700 kHz); exceptionally for leukemia cells, the frequency range was extended to 50 kHz ≤f (t )≤1550 kHz (i.e., f c =800 kHz and Δ f X =750 kHz) in the conductivity range of 60 mS/m≤σ ≤80 mS/cm because f X in this σ -range has been found to be lower than 100 kHz, and we were unable to observe the DEP U-turns in the range of 100 kHz ≤f (t )≤1500 kHz. Both of these frequency sets satisfy the WBL condition of Δ f /f m , f (t )/f m <10 −5 as before.
Each time we measured the leaving times of cells dispersed in a suspension, we looked for an appropriate spot at which a few cells having a similar size could simultaneously experience the FM-DEP above the substrate, and the microelectrode tips were placed at the measurable position using the micromanipulator. We continued such scanning inside the cell suspensions until the FM-DEP trajectories of 10 cells were collected in total at a couple of appropriate positions. For each kind of cell, the measurement of 10 cells was repeated twice using different drops of the same cell suspension. As mentioned, it is indispensable for the implementation of the FM-DEP measurement at each spot to suppress the electrically induced solvent flows as much as possible. Hence, we traced only two cycles of the U-turn path so that the duration time of applying the electric field could be adjusted to be less than 10 s, and, correspondingly, the leaving time of each cell is given as the average of each trajectory, including the two U-turns. The mean leaving time \(\overline {\Delta t_{n}}\) of each cell suspension is thus obtained from averaging the leaving times of 20 cells. Particularly for human RB cells, we further averaged three sets of the mean crossover frequencies obtained for three RB cell suspensions of three human beings, supposing that cells of the same species are similar in C m en σ cyt as well as in R . The two-step averaging of Δ t n will be denoted by \(\left <\overline {\Delta t_{n}}\right>\). Substituting into Eq. (10) the experimental data of either \(\overline {\Delta t_{n}}\) or \(\left <\overline {\Delta t_{n}}\right>\), the mean crossover frequency <f X > was obtained.
Figure 6 shows the σ uit -dependencies of <f X > measured for the above three kinds of biological cells using the FM-DEP method. The solid lines in Fig. 6 depict the best-fit results of Eq. (15). We evaluated C m en σ cyt from the best fitting of Eq. (15) into which the observed radii (R obs ) were inserted. Table 1 lists the fitting results of C m en σ cyt , where we used the observed radii of 10 μ m ≤ 2R obs ≤ 15 μ m for TL and BL cells, and 7.5 μ m ≤ 2R obs ≤ 10 μ m for RB cells in evaluating C m . It is to be noted from Table 1 that different species have different membrane capacitances, which are in good agreement with those reported in the literature [40–47]; the C m values of RB cells with stationary whole blood samples from normal (healthy) donors are in excellent agreement with our value [46, 47], but are substantially higher than those of washed RB cells in isotonic buffered saline as noted in [47]. The best-fit results simultaneously provided cytoplasmic conductivities, which were consistently similar as seen from Table 1, but were slightly lower than the range of previous reports that 0.2 S/m ≤σ cyt ≤1 S/m [40, 45, 48–51]. These results support that the FM-DEP method retains the practical reliability needed for the treatment of living cells.

Conductivity dependences of crossover frequencies. Mean crossover frequency, <f X >, of TL cells (blue triangles), BL cells (green diamonds), and RB cells (red circles) varying with increase of solution conductivity σ uit . The best-fit results of Eq. (15) are delineated by the solid lines
Conclusies
Our theoretical treatment of the FM-DEP has mainly focused on the WBL condition. In this limit, we have proved theoretically that the direction of the FM-DEP force switches each time when the instantaneous frequency of the FM wave traverses the crossover frequency, thereby implying the periodic U-turns of micro/nanoparticles that undergo the FM-DEP. Two kinds of experiment have demonstrated the accuracy and reliability of f X obtained from the observed trajectories of MLVs and cells using our formulation of the FM-DEP (Eqs. (9) and (11)):While the f X evaluated from the FM-DEP of a single MLV coincides with that obtained from the force measurement of the same MLV experiencing AC-DEP, the conductivity dependencies of f X provide the membrane capacitances of various cells that are in close agreement with the literature values. In other words, it has been validated theoretically and experimentally that the FM-DEP in the WBL limit can be mimicked by the time-varying AC-DEP induced by the AC wave with its frequency changing continuously according to the periodic function of f (t ). The simple view applies to other electrokinetics, including the twDEP and the electrorotation by applying the FM wave that has the spatial dependence of the phase as well as the magnitude. The AC- and FM-DEPs are associated with the real part of the dielectric spectra (or the CM factor), whereas the electrokinetics due to the spatial gradient of the phase reflect the imaginary part of the CM factor as mentioned before. Therefore, the application of the FM wave to either twDEP or electrorotation will be required for completing the dielectric characterization (the dielectric spectroscopy, in general) using the electrokinetics.
We have treated microparticles such as MLVs and cells for the precise tracking of particle trajectories. In these experiments, sedimented particles as well as floating ones have been observed; we need to increase the magnitude of electric field for inducing the DEP of the sedimented particles which are likely to be aggregated. Accordingly, we have used the plug-in system for applying the FM wave to a targeted particle floating above the substrate.
It is promising to further develop the FM-DEP method for smaller particles with their sizes of submicron to nanoscale, such as dispersed carbon nanotubes, thereby opening up the possibility of real-time spectroscopy using the FM-DEP as described below. When we apply the FM wave to the smaller colloids using the on-chip systems whose electrode configuration is designed to create a constant gradient of the applied electric field, the time-varying velocity vector v (t ) of the FM-DEP caused by the time dependence of the FM-DEP force is ascribed to the variation in χ (f ) (or the real part of the CM factor):it is found from Eqs. (9) and (12) that
$$ \boldsymbol{v}(t)=\frac{\nabla\boldsymbol{A}^{2}_{\text{RMS}}}{6\pi\eta R}\chi\{f(t)\}. $$ (16)Hence, measuring the velocity vector v (t ) of a submicron to nanoparticle could provide the frequency dependence of the real part of the CM factor directly, which would be nothing but the electrokinetic FM spectroscopy.
Afkortingen
- AC:
-
Alternating current
- BL:
-
B cell leukemia
- CM:
-
Clausius-Mossotti
- DEP:
-
Dielectrophoresis
- DOPC:
-
1,2-Dioleoyl-sn-glycero-3-phosphatidylcholine
- FM:
-
Frequency modulated
- MLV:
-
Multilamellar vesicle
- RB:
-
Red blood
- RMS:
-
Root mean squared
- TL:
-
T cell leukemia
- twDEP:
-
Traveling wave dielectrophoresis
- WBL:
-
Wide band limit
Nanomaterialen
- Frequentie- en fasemeting
- IoT en de impact van 5G beoordelen
- Effect van gouden nanodeeltjesdistributie in TiO2 op de optische en elektrische kenmerken van kleurstofgevoelige zonnecellen
- Synthese van ZnO-nanokristallen en toepassing in omgekeerde polymeerzonnecellen
- De structurele, elektronische en magnetische eigenschappen van Ag n V-clusters (n = 1–12) onderzoeken
- De fabricage en zeer efficiënte elektromagnetische golfabsorptieprestaties van CoFe/C Core-Shell gestructureerde nanocomposieten
- Vervaardiging en karakterisering van ZnO-nanoclips door het door polyol gemedieerde proces
- Invloed van Ag-nanodeeltjes met verschillende afmetingen en concentraties ingebed in een TiO2-compacte laag op de conversie-efficiëntie van perovskiet-zonnecellen
- Invloed van het substraat op de LSP-koppelingsgolflengte en sterkte
- Robots, cobots en automatisering:zorgt de pandemie voor een golf van verandering in de productie?
- 4 soorten voorraadcontrolesystemen:eeuwigdurende versus periodieke voorraadcontrole en de voorraadbeheersystemen die ze ondersteunen