


Neerwaartse regulatie van microRNA-342-5p of opwaartse regulatie van Wnt3a remt angiogenese en handhaaft atherosclerotische plaque-stabiliteit bij atherosclerose-muizen
Abstract
Bewijs heeft aangetoond dat microRNA-342-5p (miR-342-5p) betrokken is bij atherosclerose (AS), maar er is weinig bekend over de intrinsieke regulerende mechanismen ervan. Hier wilden we het effect onderzoeken van miR-342-5p gericht op Wnt3a op de vorming van kwetsbare plaques en angiogenese van AS. ApoE −/− muizen werden 16 w gevoed met vetrijk voer om het AS-kwetsbare plaquemodel te repliceren. miR-342-5p- en Wnt3a-expressie in aortaweefsels van AS werden gedetecteerd. De doelrelatie tussen miR-342-5p en Wnt3a werd geverifieerd. Bovendien, ApoE −/− muizen werden geïnjecteerd met miR-342-5p antagomir en overexpressie-Wnt3a-vector om hun functies in serumlipideniveaus, inflammatoire en oxidatieve stressgerelateerde cytokines, aorta-plaquestabiliteit en angiogenese in plaque van AS-muizen te testen. miR-342-5p-expressie was verbeterd en Wnt3a-expressie werd afgebroken in aortaweefsels van AS-muizen en miR-342-5p was direct gericht op Wnt3a. Opwaartse regulering van Wnt3a of neerwaartse regulering van miR-342-5p verminderde het lipidegehalte in het bloed, ontstekings- en oxidatieve stressniveaus, de kwetsbaarheid van aortaweefselplaque en remde angiogenese in aortaplaque van AS-muizen. Functionele studies tonen aan dat het uitputten van miR-342-5p plaque in aortaweefsel kan stabiliseren en angiogenese in plaque bij AS-muizen kan verminderen door Wnt3a te herstellen.
Inleiding
Atherosclerose (AS) is een leeftijdsgebonden slagaderziekte die wordt gekenmerkt door de verdikking, stenose, verharding en de vorming van atherosclerotische plaques van de slagaders [1]. Het is de belangrijkste doodsoorzaak en morbiditeit in ontwikkelde landen [2]. Histopathologische studies van menselijke atherosclerotische laesies hebben aangetoond dat plaque-ontwikkeling en -ruptuur worden gekenmerkt door lipide/necrotische kernexpansie, vermindering van het aantal gladde spiercellen, macrofaaginfiltratie en verlaagd collageengehalte [3]. De belangrijkste cellulaire elementen van AS omvatten hyperlipidemie, vorming van schuimcellen, differentiatie in macrofagen, rekrutering van monocyten en geïnduceerde ontsteking [4]. Hoewel veel geneesmiddelen voor de behandeling van AS op grote schaal in de kliniek zijn gebruikt, lopen sommige subgroepen van patiënten nog steeds een hoog risico op een myocardinfarct, myocardischemie, hartfalen en beroerte [5]. Daarom kan verdere verkenning van potentiële moleculaire mechanismen meer bewijs leveren voor AS-behandeling.
Een enkel microRNA (miRNA) kan tegelijkertijd meerdere gendoelen reguleren [6]. Van miR-342-5p is onderzocht dat het aanwezig is in het ingeprinte 14q32 miRNA-cluster, werkt als een innovatief Notch-stroomafwaarts molecuul [7] en moduleert meerdere angiogene routes, zoals transformerende groeifactor-β-signalering en vasculaire endotheliale groeifactor [8]. Gerelateerde immunomodulerende microRNA's, zoals miR-342-5p, hebben meerdere belangrijke rollen bij het reguleren van de progressie van atherosclerose [9]. Bovendien is gesuggereerd dat sommige miRNA's betrokken zijn bij de resolutie van AS, zoals miR-155 en miR-217 [10, 11]. Een studie heeft gemeld dat miR-342-5p fungeert als een nieuwe modulator voor macrofaagactivering in AS [12]. Een andere studie heeft aangetoond dat miR-342-5p afkomstig van macrofagen AS faciliteert en de inflammatoire stimulatie van macrofagen verhoogt [13]. Qu et al. hebben ontdekt dat Wnt3a-expressie negatief wordt gemoduleerd door miR-342-5p in anorectale malformaties [14], wat aangeeft dat er een doelrelatie is tussen miR-342-5p en Wnt3a. Wnt-signalering speelt een essentiële rol tijdens embryogenese voor de modulatie van celpolariteit, celproliferatie, asvorming en apoptose [15]. Wnt3a, een sleutelcomponent van het mesoderm-gen, speelt een cruciale rol in de embryonale ontwikkeling [16]. Er is gepresenteerd dat epigenoom-geleide analyse van het transcriptoom van plaque-macrofagen tijdens AS-regressie activering van de Wnt-signaleringsroute onthult [17]. Bovendien heeft een studie gerapporteerd dat Wnt3a adhesie en migratie van vasculaire gladde spiercellen moduleert die bijdragen aan de pathogenese van AS en restenose [18]. Daarom onderzocht deze studie voor het eerst het effect van miR-342-5p gericht op Wnt3a op de vorming van kwetsbare plaques en angiogenese van AS.
Materialen en methoden
Ethische verklaring
Dieren werden humaan behandeld met behulp van goedgekeurde procedures in overeenstemming met de aanbevelingen in de gids voor de zorg en het gebruik van proefdieren van de National Institutes of Health. Het protocol is goedgekeurd door de Institutional Animal Care and Use Committee van het Qinghai Provincial People's Hospital (ethisch nummer:201870726).
Experimentele dieren
Mannelijke ApoE −/− muizen en C57BL/6J-muizen (specifieke pathogeenvrije graad) van 8 weken oud waren verkrijgbaar bij Beijing Vital Laboratory Animal Technology (Beijing, China). Muizen (5-6 muizen in een kooi) werden gehuisvest met een dag-/nachtcyclus van 12 uur/12 uur met onbeperkte toegang tot voedsel en water.
Vaststelling van muizenmodellen van AS
ApoE −/− muizen werden 16 w gevoed met vetrijk voer om het AS-kwetsbare plaquemodel vast te stellen. C57BL/6 J-muizen werden gebruikt als de normale groep met natuurlijke drank en voedsel. ApoE −/− muizen hadden glanzend licht haar en haaruitval in de rug na 12 w. De aortaboog en brachiocephalische slagader van 3 modelmuizen werden ontleed om hematoxyline-eosine (HE) kleuring uit te voeren, en er was geen significante plaque-afzettingen op de intima. Nog eens 3 gemodelleerde muizen werden na 4 w opnieuw geïdentificeerd en HE-kleuring toonde aan dat er duidelijke plaque-afzettingen waren op de intima van de aortaboog, wat wijst op succes van het opzetten van het model.
Muizen groeperen en behandelen
ApoE
−/−
muizen met AS-kwetsbare plaque werden verdeeld in 6 groepen met 12 muizen in elke groep:AS-groep, negatieve controle (NC) groep (geïnjecteerd met normale zoutoplossing in ApoE
−/−
muizen), miR-342-5p agomir-groep (geïnjecteerd met miR-342-5p agomir om miR-342-5p-expressie tot overexpressie te brengen in ApoE
−/−
muizen), miR-342-5p antagomir-groep (geïnjecteerd met miR-342-5p antagomir om miR-342-5p-expressie in ApoE
−/−
te verminderen muizen), overexpressie (oe)-Wnt3a-groep (geïnjecteerd met oe-Wnt3a-vector om Wnt3a-expressie in ApoE
−/−
te verhogen muizen) en miR-342-5p agomir + oe-Wnt3a-groep (geïnjecteerd met miR-342-5p agomir en oe-Wnt3a vector om de expressie van miR-342-5p en Wnt3a in ApoE
−/−
In het voorlopige experiment, ApoE −/− muizen met AS werden geïnjecteerd met 10 mg/kg, 20 mg/kg, 40 mg/kg miR-342-5p agomir, miR-342-5p antagomir of oe-Wnt3a vector (eens per twee weken; 4 keer in totaal). Vervolgens werden de expressieniveaus van β-catenine gedetecteerd door kwantitatieve polymerasekettingreactie met omgekeerde transcriptie (RT-qPCR).
Monsterverzameling en behandeling
Vóór de bemonstering werden muizen 12 uur gevast en verdoofd door etherinhalatie en werden bloedmonsters verzameld uit oogbollen. De borstkas van muizen werd geopend, de thoracale aorta werd losgekoppeld van het uiteinde van de abdominale aorta en het hele vat werd verwijderd. Na reiniging met RNA-vrije fosfaatgebufferde zoutoplossing (PBS) werden de weefsels ingebed voor HE-kleuring, olie-rood O, Sirius-rood-kleuring en immunohistochemische kleuring. Sommige vaatweefsels werden bewaard bij -80 °C voor RT-qPCR en Western-blot.
Detectie van bloedlipideniveaus
De automatische biochemische analysator (Roche, Basel, Zwitserland) werd gebruikt om totaal cholesterol (TC), triglyceride (TG), lipoproteïne-cholesterol met lage dichtheid (LDL-C) en lipoproteïne-cholesterol met hoge dichtheid (HDL-C) in serum te detecteren. De detectie werd geïmplementeerd volgens de specificatie van de kits (NanJing JianCheng Bioengineering Institute, Nanjing, China).
Enzym-Linked Immunosorbent Assay (ELISA)
Bepaling van het serumcytokinegehalte:er werden commerciële interleukine (IL)-5, IL-12p70, tumornecrosefactor-alfa (TNF-α) en interferon (IFN)-γ ELISA-kits gebruikt. Ten slotte werd de optische dichtheidswaarde (OD) van elk putje getest door een microplaatlezer bij 450 nm.
Bepaling van schade door oxidatieve stress:malondialdeyde (MDA)-gehalte en superoxide-dismutase (SOD)-activiteit in serum werden getest door MDA-kit (OD-waarde werd getest door spectrofotometer bij 532 nm) en SOD-kit (OD-waarde werd bepaald door een microplaatlezer bij 450 nm). IL-5, IL-12p70, TNF-α, IFN-γ, MDA en SOD ELISA-kits werden gekocht bij MultiSciences (Lianke) Biotechnology Corporate Limited (Hangzhou, Zhejiang, China).
HE-kleuring, olierode O-kleuring en Sirius-rode kleuring
Na fixatie en inbedding werden de monsters in opeenvolgende secties met een dikte van 4 microm gesneden. Plakjes werden van was ontdaan en gehydrateerd, gekleurd met hematoxyline en eosine, gedifferentieerd, gedehydrateerd, helder gemaakt met xyleen, gedroogd en afgesloten met neutrale gom. De kern was blauw en andere weefsels zoals cytoplasma en bindweefsel waren rood in verschillende tinten. De plaquevorming werd waargenomen met een fluorescentiemicroscoop. De arteriële wanden van HE-gekleurde secties werden onder de microscoop geselecteerd en de experimentele resultaten werden verzameld door een digitale camera. De Image Pro Plus6.0 (IPP6.0)-softwaremodule voor beeldanalyse werd gebruikt om het plaque-oppervlak van de dwarsdoorsnede van elke plak en het oppervlak van de wand en hun verhouding te berekenen.
Plakjes met 4-5 μm werden gekozen voor olierode O-kleuring. De plakjes werden gedurende 20 minuten bij overmatige temperatuur gedroogd en gedurende 5 minuten met 100% isopropanol geïncubeerd. Vervolgens werden de plakjes 8 minuten geïncubeerd met 0,5% olie-rood O-kleuringsoplossing in een oven van 60 ° C, 3 minuten gewassen in 85% isopropanol, 1 minuut geverfd met hematoxyline, schoongemaakt en afgesloten. De resultaten van olierood O-kleuring suggereerden dat het lipide rood of oranje was en de kern lichtblauw. IPP6.0-software werd gebruikt om het vetgebied en het plaquegebied in de plaque van het weefselplakje te berekenen. Het lipidegehalte = olierood O-positief kleuringsgebied/plaquegebied × 100%.
Sirius rode kleuring:plakjes werden van was ontdaan en gehydrateerd, gedurende 10 minuten geverfd met celestineblauwe kleuroplossing, met Sirius rode kleuroplossing gedurende 20 minuten en gedurende 10 minuten tegengekleurd met hematoxyline. Ten slotte werden plakjes gedehydrateerd met gradiënt-ethanol, geklaard met xyleen en afgesloten met neutrale gom. Het collageengebied in de plaque van het weefselplakje werd berekend met IPP6.0-software. Het collageengebied = Sirius rood positief kleuringsgebied/plaquegebied × 100%. Het percentage lipiden en collageen in het plaquegebied werd berekend.
Hematoxyline, eosine en Sirius-kleurstof waren verkrijgbaar bij China Pharmaceutical Group Shanghai Chemical Reagent Co. Ltd. (Shanghai, China). Het olierode O-poeder werd gekocht bij Sigma-Aldrich Chemical Company (St Louis, MO, VS).
Kwantitatieve polymerasekettingreactie met omgekeerde transcriptie (RT-qPCR)
De aortaweefsels werden toegevoegd aan totaal RNA-extractiereagens Trizol (Invitrogen, Carlsbad, Californië, VS) en vervolgens gehomogeniseerd om het totale RNA en samengesteld complementair DNA te extraheren. De primers werden allemaal verwezen naar de sequentie geleverd door Genbank, ontworpen door Primer 5.0 en gesynthetiseerd door Shanghai Sangon Biotechnology Co. Ltd. MiR-342-5p:forward:5'-CGGAGGGGTGCTATCTGTGATTGAG-3', reverse primers waren kit universele primers (Qiagen bedrijf, Hilden, Duitsland); Wnt3a:vooruit:5′-AGGTAAGCTACTCCCTCAACTA-3′, achteruit:5′-CTGAAGCACCCTCTCATGTATC-3′; β-actine:vooruit:5'-GCACCACACCTTCTACAATGAGC -3', achteruit:5'-TCGTTGCCAATAGTGATGACC-3'; β-catenine:vooruit:5′-TCAAGAGAGCAAGCTCATCATTCT-3′, achteruit:5′-CACCTTCAGCACTCTGCTTGTG-3′. Na de reactie werd de drempelcyclus (Ct) met de computer geanalyseerd. De relatieve verhouding van miR-342-5p tot U6 werd gebruikt als uitdrukking, de relatieve verhouding van Wnt3a tot β-actine werd gebruikt als uitdrukking en de relatieve verhouding werd berekend met 2 −ΔΔCt methode.
Western Blot-analyse
Het totale eiwit werd geabstraheerd uit de aortaweefsels. De eiwitconcentratie werd gemeten met de bicinchoninezuurmethode. Polyacrylamidegelelektroforese werd uitgevoerd. Vervolgens werd eiwit overgebracht naar het polyvinylideenfluoridemembraan en werd de doelband verkregen. Het membraan werd gedurende 1 uur afgesloten in 5% magere melk, toegevoegd met primaire antilichamen Wnt3a (1:500), β-catenine (1:1000, Santa Cruz Biotechnology, Inc., Santa Cruz, CA, VS), CD34 (1 :2500, Abcam, MA, VS) en β-actine (1:2000, Beyotime Biotechnology Co., Shanghai, China) 's nachts bij 4 ° C. Het membraan werd 10 min × 3 keer gewassen met Tris-gebufferde zoutoplossing met Tween 20 (pH = 7,5, 10 mmol/L Tris-HCl, 100 mmol/L NaCl en 0,2% Tween-20) en daarna toegevoegd met secundair antilichaam. (1:1000, ZSGB-Bio, Peking, China) gedurende 2 uur. ImageJ-software is gebruikt om de grijswaarde van banden te beoordelen en eiwitexpressie te kwantificeren.
Immunohistochemische kleuring
Plakjes van 4-5 m werden op de glaasjes geplaatst die waren gecoat met 100 mg / L polylysine en gefixeerd met aceton. Endogeen peroxidase werd geblokkeerd door runderserumalbumine. De weefsels werden gedruppeld met MOMA-2-antilichaam (1:200), α-SMA (1:200) en CD34 (1:200, Abcam Inc., Cambridge, MA, VS) en toegevoegd met secundaire antilichaam-werkoplossing ( 1:1000). De weefsels werden ontwikkeld met diaminobenzidine, tegengekleurd met hematoxyline (1 min), gedehydrateerd, geklaard, afgesloten en onder de microscoop bekeken. Voor elke immunohistochemische sectie werden drie verschillende gezichtsvelden geselecteerd. IPP6.0-software werd uitgevoerd voor kwantitatieve analyse. Positieve immunohistochemische kleuring van respectievelijk MOMA-2 en α-SMA geeft aan dat macrofagen en gladde spiercellen zich voornamelijk in het cytoplasma bevinden, dat geel tot bruin is. De percentages macrofagen en gladde spiercellen werden afzonderlijk berekend, die werden gecombineerd met het percentage lipiden en collageen in de plaque om de plaquekwetsbaarheidsindex te berekenen. De kwetsbaarheidsindex van plaque = (positief percentage macrofagen + positief percentage lipiden)/(positief percentage collageen + positief percentage gladde spiercellen) [20]. Microvatdichtheid (MVD) werd bepaald door meting van CD34-expressie en gekwantificeerd als het aantal microvaten/mm 2 .
Dual Luciferase Reporter Gene Assay
Het doelgen van miR-342-5p werd geanalyseerd door een biologische voorspellingswebsite (//www.microRNA.org). Dubbele luciferase-reportergentest werd gebruikt om te verifiëren of Wnt3a het doelwitgen was van miR-342-5p. De wildtype of mutante sequentie van Wnt3a 3'-niet-vertaalde regio (3'-UTR) werd gekloneerd in de GP-miRGLO-vector (GenePharma, Shanghai, China). De reporter (0,5 μg) en 1, 10 of 100 pM miR-342-5p agomir werden gedurende 48 uur getransfecteerd in aorta-endotheelcellen van de muis (nr. 506, MingzhouBio, Ningbo, China) om de luciferase-activiteit te testen met behulp van een dubbel luciferase-assaysysteem (Promega, WI, VS).
Statistische analyse
Alle gegevens werden geïnterpreteerd door SPSS 21.0-software (IBM Corp. Armonk, NY, VS). Meetgegevens werden aangegeven als gemiddelde ± standaarddeviatie. Verschillen tussen twee groepen werden geformuleerd door t -test, terwijl die tussen meerdere groepen door eenrichtingsanalyse van variantie (ANOVA) gevolgd door Tukey's meervoudige vergelijkingstest. Statistische significantie werd vastgesteld door P waarde < 0,05.
Resultaten
miR-342-5p-verhogingen en Wnt3a-verlagingen in aortaweefsels van ApoE −/− muizen en miR-342-5p richten zich direct op Wnt3a
MicroRNA (miRNA) doelgenen zijn geassocieerd met atherosclerose-gerelateerde functies. miR-342-5p, Wnt3a en β-catenine werden getest in aortaweefsels van AS-modelmuizen met RT-qPCR en Western-blot-assay. Er werd onthuld dat in verhouding tot de normale groep miR-342-5p verhoogd was, terwijl Wnt3a en β-catenine verlaagd waren in de AS-groep (beide P < 0.05). In vergelijking met de NC-groep was miR-342-5p verbeterd en waren Wnt3a en β-catenine verlaagd in de miR-342-5p agomir-groep (beide P <-0,05), terwijl miR-342-5p was verlaagd, waren Wnt3a en β-catenine verhoogd in de miR-342-5p antagomir-groep (beide P < 0.05). Wnt3a- en β-catenine-expressie waren verhoogd in de oe-Wnt3a-groep ten opzichte van de NC-groep (beide P < 0.05). Vergeleken met de miR-342-5p agomir-groep, waren Wnt3a- en β-catenine-expressie verhoogd in de miR-342-5p agomir + oe-Wnt3a-groep (P <-0.05) (Fig. 1A-D). Bovendien werd in het voorlopige experiment β-catenine-expressie onder de behandeling van verschillende concentraties van miR-342-5p agomir, miR-342-5p antagomir en oe-Wnt3a getest en de resultaten werden getoond (aanvullend bestand 1:Fig. S1 ) hoe hoger de miR-342-5p agomir-concentratie, hoe lager de β-catenine-expressie; hoe hoger de miR-342-5p antagomir-concentratie, hoe hoger de β-catenine-expressie; en hoe hoger de oe-Wnt3a-concentratie, hoe hoger de β-catenine-expressie.
miR-342-5p neemt toe en Wnt3a neemt af in aortaweefsels van ApoE −/− muizen en miR-342-5p richten zich direct op Wnt3a. A Expressie van miR-342-5p in aortaweefsel van muizen gedetecteerd met RT-qPCR. B Expressie van Wnt3a-mRNA in aortaweefsel van muizen gedetecteerd met RT-qPCR (n = 12). C , D Wnt3a- en β-catenine-eiwitexpressie in aortaweefsels van muizen getest met Western-blot-analyse (n = 12). E Bindingsplaats van miR-342-5p binnen Wnt3a 3'-UTR. V miR-342-5p agomir verlaagde dosisafhankelijk de relatieve activiteit in cellen die waren getransfecteerd met Wnt3a 3′-UTR (N = 3). G Relatieve activiteit van luciferase in cellen met wildtype en mutant Wnt3a 3'-UTR na transfectie met miR-342-5p agomir of scramble (N = 3). *P < 0.05 vs. de normale groep, # P < 0,05 vs. de NC-groep. &P <-0,05 versus de miR-342-5p agomir-groep. Meetgegevens werden aangegeven als gemiddelde ± standaarddeviatie. Vergelijkingen tussen twee groepen zijn gemaakt door t -test, terwijl vergelijkingen tussen meerdere groepen werden beoordeeld door eenrichtings-ANOVA gevolgd door Tukey's meervoudige vergelijkingstest. AS, atherosclerose; NC, negatieve controle
miRNA's zouden de translatie van specifieke genen kunnen remmen door te binden aan hun boodschapper-RNA 3'UTR. Bioinformatica-website voorspelde dat er een doelrelatie was tussen miR-342-5p en Wnt3a (Fig. 1E). Dubbele luciferase-reportergen-assay meldde dat in met Wnt3a 3'UTR-vector getransfecteerde aorta-endotheelcellen van muizen de renilla / vuurvliegwaarde van luciferase dosisafhankelijk werd verlaagd door transfectie met miR-342-5p agomir, met een significante afname van 10 naar 100 pM miR-342-5p agomir en een afname van 64% trad op bij 100 pM miR-342-5p agomir-groep in vergelijking met de NC-groep. Dit duidde op de aanwezigheid van een miR-342-5p-doellocatie in de Wnt3a 3'UTR. De renilla / vuurvliegwaarde van luciferase-activiteit werd echter niet beïnvloed in de Wnt3a-mutatiegroep (Fig. 1F, G). Het kon dus worden bevestigd dat Wnt3a een direct doelwitgen was van miR-342-5p en dat miR-342-5p/Wnt3a de progressie van AS zou kunnen reguleren.
Effecten van omhoog gereguleerde Wnt3a of omlaag gereguleerde miR-342-5p op de lipideniveaus in ApoE −/− Muizen
Bovendien, om te onderzoeken of miR-342-5p-targeting en regulering van de Wnt3a-signaleringsroute de lipideniveaus van AS-muizen zou beïnvloeden, werd een automatische biochemische analysator gebruikt om de verandering van lipideniveaus te observeren. De resultaten onthulden dat (Fig. 2A-D) in tegenstelling tot de normale groep, het TC-, TG- en LDL-C-gehalte was verhoogd en het HDL-C-gehalte was verlaagd in de AS-groep (alle P < 0.05). Vergeleken met de NC-groep was het TC-, TG- en LDL-C-gehalte verhoogd en het HDL-C-gehalte verlaagd in de miR-342-5p agomir-groep (alle P <-0,05), terwijl het TC-, TG- en LDL-C-gehalte was verlaagd en het HDL-C-gehalte was verhoogd in de miR-342-5p antagomir-groep en de oe-Wnt3a-groep (alle P < 0.05). Met betrekking tot de miR-342-5p agomir-groep was het TC-, TG- en LDL-C-gehalte verlaagd en het HDL-C-gehalte verhoogd in de miR-342-5p agomir + oe-Wnt3a-groep (alle P < 0.05). Deze resultaten suggereerden dat miR-342-5p en Wnt3a het bloedlipideniveau van AS-muizen reguleerden, en illustreerden verder de gerichte regulatierelatie tussen miR-342-5p en Wnt3a. Overexpressie van Wnt3a zou tot overexpressie gebrachte miR-342-5p-geïnduceerde effecten op AS-muizen omkeren.
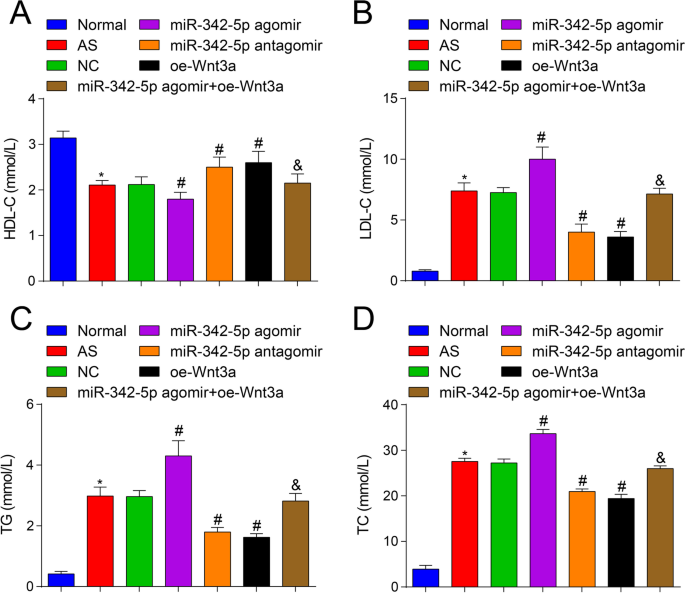
Effecten van opwaarts gereguleerd Wnt3a of neerwaarts gereguleerd miR-342-5p op de lipideniveaus in ApoE −/− muizen. A Vergelijking van HDL-C-gehaltes in serum van muizengroep. B Vergelijking van LDL-C-gehaltes in serum van muizengroep. C Vergelijking van TG-gehaltes in serum van muizengroep. D Vergelijking van TC-gehaltes in serum van muizengroep. n = 12. *P < 0.05 vs. de normale groep, # P < 0,05 vs. de NC-groep. &P <-0,05 versus de miR-342-5p agomir-groep. Meetgegevens werden aangegeven als gemiddelde ± standaarddeviatie. Vergelijkingen tussen meerdere groepen werden beoordeeld door eenrichtings-ANOVA gevolgd door Tukey's meervoudige vergelijkingstest. AS, atherosclerose; NC, negatieve controle
Effecten van overexpressie van Wnt3a of lage expressie van miR-342-5p op inflammatoire en oxidatieve stressgerelateerde cytokines in serum van ApoE −/− Muizen
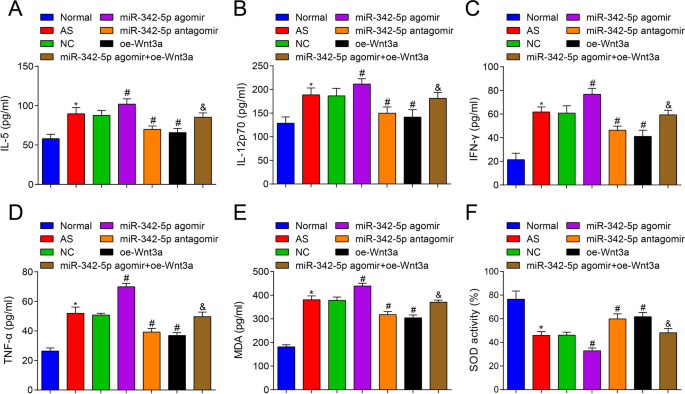
Vervolgens werd de inhoud van cytokinen in serum van AS-muizen getest met ELISA en de resultaten meldden dat (Fig. 3A-D) versus de normale groep, IL-5, IL-12p70, IFN-γ en TNF-α waren verbeterd in de AS-groep (allemaal P < 0.05). Vergeleken met de NC-groep waren IL-5-, IL-12p70-, IFN-γ- en TNF-α-gehalten verhoogd in de miR-342-5p agomir-groep (alle P <-0,05), terwijl het gehalte aan IL-5, IL-12p70, IFN-γ en TNF-α werd afgebroken in de miR-342-5p antagomir-groep en de oe-Wnt3a-groep (allemaal P < 0.05). In tegenstelling tot de miR-342-5p agomir-groep, waren de IL-5-, IL-12p70-, IFN-γ- en TNF-α-gehalten verlaagd in de miR-342-5p agomir + oe-Wnt3a-groep (alle P < 0.05). Er werd gesuggereerd dat miR-342-5p gerichte regulatie van de Wnt3a-signaleringsroute het niveau van gerelateerde cytokines in serum van AS-muizen verder regelde.
Effecten van overexpressie van Wnt3a of lage expressie van miR-342-5p op inflammatoire en oxidatieve stress-gerelateerde cytokines in serum van ApoE −/− muizen. A Vergelijking van IL-5-gehaltes in serum van muizengroep. B Vergelijking van IL-12p70-gehaltes in serum van muizengroep. C Vergelijking van IFN-γ-gehaltes in serum van muizengroep. D Vergelijking van TNF-a-gehaltes in serum van muizengroep. E, Vergelijking van MDA-gehalte in serum van muizengroep. V Vergelijking van SOD-activiteit in serum van muizengroep. n = 12. *P < 0.05 vs. de normale groep, # P < 0,05 vs. de NC-groep. &P <-0,05 versus de miR-342-5p agomir-groep. Meetgegevens werden aangegeven als gemiddelde ± standaarddeviatie. Vergelijkingen tussen meerdere groepen werden beoordeeld door eenrichtings-ANOVA gevolgd door Tukey's meervoudige vergelijkingstest. AS, atherosclerose; NC, negatieve controle
Verder werden het MDA-gehalte en de SOD-activiteit in serum van muizen getest, en er werd onthuld dat (Fig. 3E, F) in vergelijking met de normale groep, het MDA-gehalte was verhoogd en de SOD-activiteit was verlaagd in de AS-groep (beide P < 0.05). Vergeleken met de NC-groep was het MDA-gehalte verhoogd en de SOD-activiteit verlaagd in de miR-342-5p agomir-groep (beide P <-0,05), terwijl het MDA-gehalte was verlaagd en de SOD-activiteit was verhoogd in de miR-342-5p antagomir-groep en de oe-Wnt3a-groep (alle P < 0.05). Vergeleken met de miR-342-5p agomir-groep, was het MDA-gehalte verlaagd en was de SOD-activiteit verhoogd in de miR-342-5p agomir + oe-Wnt3a-groep (beide P < 0.05). Daarom werd een samenvatting verkregen dat het uitputten van miR-342-5p en het herstellen van Wnt3a oxidatieve stress in AS-muizen remde.
Effecten van uitputting van miR-342-5p of herstel van Wnt3a op lipiden- en collageeninhoud in aortaplaque van ApoE −/− Muizen
Om het effect van miR-342-5p gericht op Wnt3a op plaquegebied in aortaweefsel van muizen te onderzoeken, werd HE-kleuring uitgevoerd en de resultaten onthulden dat (Fig. 4A, B) behalve de normale groep, AS-plaques gevormd in alle delen van de andere groepen. In de AS-groep was het plaquegebied groot, de fibreuze kap was dunner, de lipidekern was vergroot, er verschenen meer schuimcellen en cholesterolkristalprecipitatie in de plaque, de binnenwand van de slagader en de spierlaag waren verdikt en de plaque was onstabiel. De situatie in de NC-groep was vergelijkbaar met die van de AS-groep. In de miR-342-5p antagomir en de oe-Wnt3a-groep was het plaquegebied klein, de intima van de slagader was glad en fibreuze caps hadden een klein aantal en werden dunner. Er was geen breuk maar schuimcellen in verschillende maten in de plaque. Het cholesterolkristal was asymmetrisch verdeeld en gedeeltelijk verkalkt, het aantal gladde spiercellen en collageenvezels was toegenomen en de plaque had de neiging stabiel te zijn. Vergeleken met de NC-groep was het plaque-oppervlak toegenomen en waren AS-laesies verergerd in de miR-342-5p agomir-groep (P <-0,05) terwijl het plaquegebied was afgenomen in de miR-342-5p antagomir-groep en de oe-Wnt3a-groep met verminderde AS-laesies (beide P < 0.05). Vergeleken met de miR-342-5p agomir-groep, was het plaquegebied afgenomen in de miR-342-5p agomir + oe-Wnt3a-groep (P < 0.05).
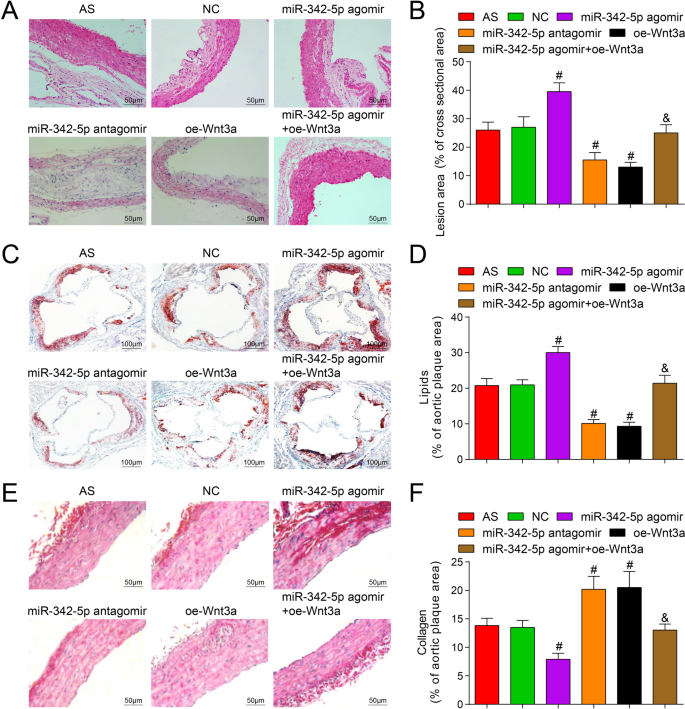
Effecten van uitputting van miR-342-5p of herstel van Wnt3a op het lipiden- en collageengehalte in aortaplaque van ApoE −/− muizen. A Resultaten van aorta HE-kleuring bij muizen (schaalbalk:50 m). B Vergelijking van het gebied van de aortaplaque in elke groep muizen. C Resultaten van aorta Oil Red O-kleuring in alle groepen muizen (schaalbalk:100 m). D Vergelijking van het lipidegehalte in aortaweefsel van muizen. E Resultaten van aorta Sirius rode kleuring in elke groep muizen (schaalbalk:50 m). V Vergelijking van collageeninhoud in aortaweefsels van muizen. n = 12. # P < 0,05 vs. de NC-groep. &P <-0,05 versus de miR-342-5p agomir-groep. Meetgegevens werden aangegeven als gemiddelde ± standaarddeviatie. Vergelijkingen tussen meerdere groepen werden beoordeeld door eenrichtings-ANOVA gevolgd door Tukey's meervoudige vergelijkingstest. AS, atherosclerose; NC, negatieve controle
Olierood O-kleuring en Sirius-roodkleuring werden toegepast om het effect van miR-342-5p-gerichte Wnt3a op het lipidegehalte en collageengehalte in plaque van aortaweefsel van muizen te detecteren, en de resultaten toonden aan dat (Fig. 4C-F) Olierood O-kleuring vertoonde rood vet en blauwe kern, terwijl Sirius rode kleuring rode collageenvezels en blauwe kern vertoonde. Vergeleken met de NC-groep was het lipidegehalte verhoogd en het collageengehalte verlaagd in de miR-342-5p agomir-groep, evenals het lipidegehalte werd verlaagd en het collageengehalte werd geaccumuleerd in de miR-342-5p antagomir-groep en de oe-Wnt3a groep (alle P < 0.05). Met betrekking tot de miR-342-5p agomir-groep was het lipidengehalte verlaagd en het collageengehalte verhoogd in de miR-342-5p agomir + oe-Wnt3a-groep (beide P < 0.05). De experimentele resultaten illustreerden volledig dat miR-342-5p gerichte regulatie van Wnt3a-signaleringsroute een regulerend effect had op het lipide- en collageengehalte in aortaplaque van AS-muizen.
Effecten van naar beneden gereguleerde miR-342-5p of naar boven gereguleerde Wnt3a op macrofagen en gladde spiercellen in aortaplaque van ApoE −/− Muizen
De mate van AS is recht evenredig met het gehalte aan mononucleaire macrofagen [21]. VSMC's zijn de belangrijkste cellen in de middelste laag van slagaders en zijn essentieel voor het behoud van de integriteit van de slagaderwand. VSMC's zijn betrokken bij de reconstructie van de arteriële wand en spelen een belangrijke rol bij AS in verschillende stadia [22]. α-SMA is een specifieke marker van gladde spiercellen [23]. In deze studie werd een macrofaagmarkerantilichaam (MOMA-2) gebruikt om macrofagen te labelen, en immunohistochemie werd toegepast om respectievelijk MOMA-2- en α-SMA-expressie te detecteren.
Onder de microscoop geeft positieve immunohistochemische kleuring van respectievelijk MOMA-2 en α-SMA aan dat macrofagen en gladde spiercellen zich voornamelijk in het cytoplasma bevinden, dat geel tot bruin is. MOMA-2 immune positive indicated that macrophages was mainly located in the cytoplasm with yellow to brown. Determined by immunohistochemistry, it was manifested that versus the NC group, percentage of plaque macrophages (MAMO-2) positive staining was raised and percentage of positive smooth muscle cells was decreased in the miR-342-5p agomir (both P < 0.05). Percentage of plaque macrophages (MAMO-2) positive staining was reduced and percentage of positive smooth muscle cells was increased in the miR-342-5p antagomir group and the oe-Wnt3a group (all P < 0.05). In comparison with the miR-342-5p agomir group, percentage of plaque macrophages (MAMO-2) positive staining was depressed and percentage of positive smooth muscle cells was raised in the miR-342-5p agomir + oe-Wnt3a group (both P < 0.05) (Fig. 5A–D). It was implied that miR-342-5p targeted regulation of Wnt3a signaling pathway could regulate aggregation of macrophages and smooth muscle cells in arterial tissue plaques of AS mice.

Effects of high expression of Wnt3a or poor expression of miR-342-5p on aortic plaque vulnerability of ApoE −/− mice. A Immunohistochemical staining of MOMA-2 in each group of mice (scale bar:50 μm). B Quantitative analysis of figure A. C Immunohistochemical staining of α-SMA in each group of mice (scale bar:50 μm). D Quantitative analysis of figure C . E Comparison of plaque vulnerability index in aortic tissues of AS mice. n = 12. # P < 0.05 vs. the NC group. &P < 0.05 vs. the miR-342-5p agomir group. Measurement data were indicated as mean ± standard deviation. Comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s multiple comparisons test. AS, atherosclerosis; NC, negative control
Effects of High Expression of Wnt3a or Poor Expression of miR-342-5p on Aortic Plaque Vulnerability of ApoE −/− Mice
Plaque vulnerability index was calculated:(positive percentage of macrophages + positive percentage of lipids)/(positive percentage of smooth muscle cells + positive percentage of collagen). In relation to the NC group, the plaque vulnerability index was raised in the miR-342-5p agomir group (P < 0.05) and decreased in the miR-342-5p antagomir group and the oe-Wnt3a group (both P < 0.05). Versus the miR-342-5p agomir group, the plaque vulnerability index was decreased in the miR-342-5p agomir + oe-Wnt3a group (P < 0.05) (Fig. 5E). Briefly, miR-342-5p targeted regulation of Wnt3a signaling pathway-mediated vulnerability of plaques in arterial tissues of AS mice.
Effects of Low Expression of miR-342-5p or Overexpression of Wnt3a on Angiogenesis in Aortic Plaque of ApoE −/− Mice
Antibodies against endothelial cell marker CD34 can detect blood vessel density [24]. By immunohistochemistry and Western blot. Versus the NC group, MVD was heightened in the miR-342-5p agomir group and attenuated in the miR-342-5p antagomir group and the oe-Wnt3a group (all P < 0.05). In comparison with the miR-342-5p agomir group, MVD was decreased in the miR-342-5p agomir + oe-Wnt3a group (P < 0.05) (Fig. 6A–C). Collectively, miR-342-5p targeting and regulating Wnt3a signaling pathway directly participated in regulating the density of neovascularization in plaques of AS mice.

Effects of low expression of miR-342-5p or overexpression of Wnt3a on MVD in aortic plaque of ApoE −/− mice. A Immunohistochemical staining of CD34 in each group of ApoE −/− mice (scale bar:50 μm). B Comparison of MVD in aortic plaque in ApoE −/− mice. C Comparison of CD34 protein expression in ApoE −/− mice. n = 12. # P < 0.05 vs. the NC group. &P < 0.05 vs. the miR-342-5p agomir group. Measurement data were indicated as mean ± standard deviation. Comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s multiple comparisons test. AS, atherosclerosis; NC, negative control
Discussion
AS is an unpredictable disease involving forms of chronic inflammation and vascular remodeling, and is the major cause of mortality and morbidity globally [25]. A previous study has discussed that miR-342-5p is implicated in regulating the progression of AS [26]. Also, it was mirrored that Wnt pathway is involved in facilitating the occurrence and development of diabetic AS [27]. As the related mechanisms of miR-342-5p and Wnt3a in AS remain to be excavated, our study was to inquire the effect of miR-342-5p targeted Wnt3a on formation of vulnerable plaque and angiogenesis of AS.
Our study revealed that highly expressed miR-342-5p and lowly expressed Wnt3a were found in aortic tissues of AS mice. A study has presented that macrophage-derived miR-342-5p is dramatically raised in early atherosclerotic lesions in ApoE −/− mice [13]. Another study has presented that miR-342-5p is markedly elevated in atrial fibrillation patients [28]. It has been reported that Wnt3a deficiency irreversibly injures hematopoietic stem cell self-renewal and results in defects in progenitor cell differentiation [29]. A study has shown that depletion of Wnt3a leads to defective cardiac function [30]. It has been revealed that Wnt3a expression in hippocampus of Alzheimer’s disease mice is remarkably decreased [31]. Another result from our study is that Wnt3a was directly targeted by miR-342-5p in AS mice. It has been reported that miR-342-5p can target the 3'-UTR of Wnt3a and negatively regulate its expression [14].
In addition, our study has suggested that TC, TG, LDL-C, IL-5, IL-12p70, IFN-γ, TNF-α and MDA contents were increased in serum, and HDL-C content and SOD activity were decreased. In addition, plaque area, lipid content, collagen content and MVD were enhanced as well as MOMA-2 expression was raised and α-SMA expression was decreased in AS mice. IFN-γ is a soluble cytokine with many functions, including anti-fibrosis, anti-proliferation, immunomodulation, apoptosis and anti-viral activities [32]. It has been revealed that glutamine treatment markedly raises SOD activity and reduces MDA content as well as increases Wnt3a protein levels in Alzheimer’s disease [31]. A study has revealed that the plasma levels of TC, TG and LDL-C are notably elevated and HDL-C is markedly reduced in AS [33]. A study has reported that TEMPOL supplementation, which has a value in suppressing metabolic disorders and raising atherosclerotic plaque stability, enhances plaque collagen content and reduces lipid content [34]. Zhou et al. noted that OPCRR treatments dramatically reduces the serum lipid profiles including TC, TG and LDL-C as well as and raises the HDL-C, also decreases MDA content as a product of lipid peroxidation and, moreover, declines serum levels of TNF-α in AS [35]. It has been presented that atherosclerotic samples have obviously reduced expression of α-SMA [36]. A study has presented that raised MVD is found in diseased aortas and especially in ruptured atherosclerotic plaque [37]. Furthermore, our study revealed that poor expression of miR-342-5p and overexpression of Wnt3a decreased he lipid levels, cytokine contents, oxidative stress response, plaque area and lipid content as well as increased collagen content, depleted MOMA-2 expression and restored α-SMA expression in aortic tissues in AS mice. It has been suggested previously that miR-342-5p is found to be positively linked to LDL-C and TNF-α serum levels and has an inverse correlation with HDL-C in coronary artery disease (CAD) patients [12]. Another study has verified that depletion of miR-342-5p inhibits AS [13]. Additionally, an experiment has presented that low serum level of Wnt1 is related to raised TG and LDL-C in premature CAD patient [38]. In addition, a study has showed that up-regulated Wnt3a, enhanced SOD content and decreased MDA content are found in the curcumin groups in Parkinson's disease rats [39].
Conclusion
In brief, our study for the first time discovered the mechanism of miR-342-5p/Wnt3 axis in AS and revealed that depleting miR-342-5p could reduce formation of vulnerable plaque and angiogenesis in AS mice via restoring Wnt3a, which may be a potential candidate for treatment of AS (Additional file 2:Fig. S2). miR-342-5p may have a synergistic effect with other miRNAs in atherosclerotic vascular disease, but due to time and funding constraints, we did not conduct further relevant discussions, which is also a limitation of this study.
Availability of data and materials
The original contributions presented in the study are included in the article/Supplementary Material, and further inquiries can be directed to the corresponding author.
Abbreviations
- miR-342-5p:
-
MicroRNA-342-5p
- A:
-
Atherosclerosis
- α-SMA:
-
α-Smooth muscle actin
- MVD:
-
Microvessel density
- miRNA:
-
MicroRNA
- oe:
-
Overexpression
- NC:
-
Negative control
- PBS:
-
Phosphate-buffered saline
- TC:
-
Total cholesterol
- TG:
-
Triglyceride
- LDL-C:
-
Low-density lipoprotein cholesterol
- HDL-C:
-
High-density lipoprotein cholesterol
- ELISA:
-
Enzyme-linked immunosorbent assay
- IL:
-
Interleukin
- TNF-α:
-
Tumor necrosis factor alpha
- IFN:
-
Interferon
- MDA:
-
Malondialdeyde
- SOD:
-
Superoxide dismutase
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
Nanomaterialen
- Kenmerken van frequentieafwijkingen van kwartskristallen:frequentietolerantie, frequentiestabiliteit en veroudering
- Conditiebewaking verhoogt en handhaaft de efficiëntie in de papierproductie
- Cyclocarbon maken en in beeld brengen
- Grafeen in luidsprekers en oortelefoons
- Neerwaartse regulatie van microRNA-342-5p of opwaartse regulatie van Wnt3a remt angiogenese en handhaaft atherosclerotische plaque-stabiliteit bij atherosclerose-muizen
- Experimenteel onderzoek naar stabiliteit en natuurlijke convectie van TiO2-water nanovloeistof in behuizingen met verschillende rotatiehoeken
- Geminiaturiseerde rekbare en snelle lineaire supercondensatoren
- First-Principles Study on the Stability and STM Image of Borophene
- De potentiële lever-, hersen- en embryotoxiciteit van titaniumdioxide-nanodeeltjes op muizen
- Hydrothermische synthese van In2O3 nanodeeltjes hybride tweeling hexagonale schijf ZnO heterostructuren voor verbeterde fotokatalytische activiteiten en stabiliteit
- Voorbereiding van hiërarchische poreuze Silicalite-1 ingekapselde Ag NP's en zijn katalytische prestaties voor 4-nitrofenolreductie